

Coordinated metabolic responses to cyclophilin D deletion in the developing heart
- PMID: 38414851
- PMCID: PMC10897919
- DOI: 10.1016/j.isci.2024.109157
Coordinated metabolic responses to cyclophilin D deletion in the developing heart
Abstract
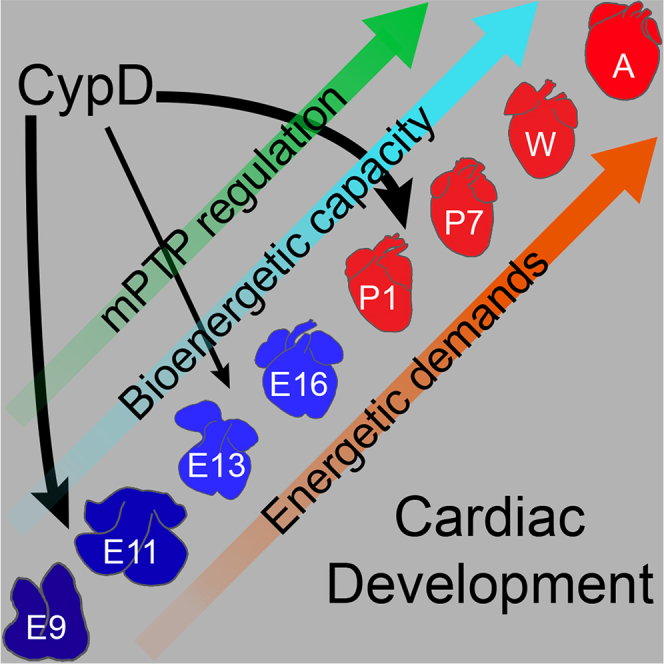
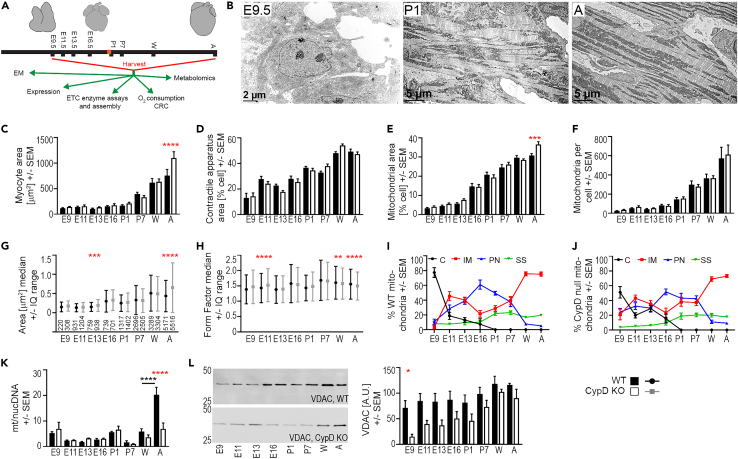
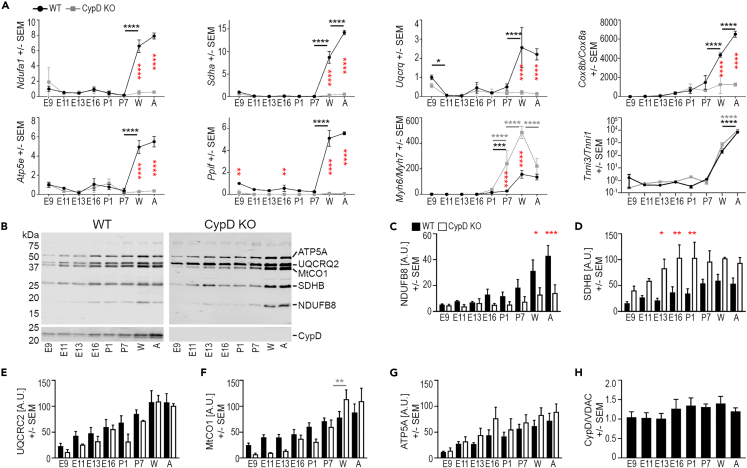
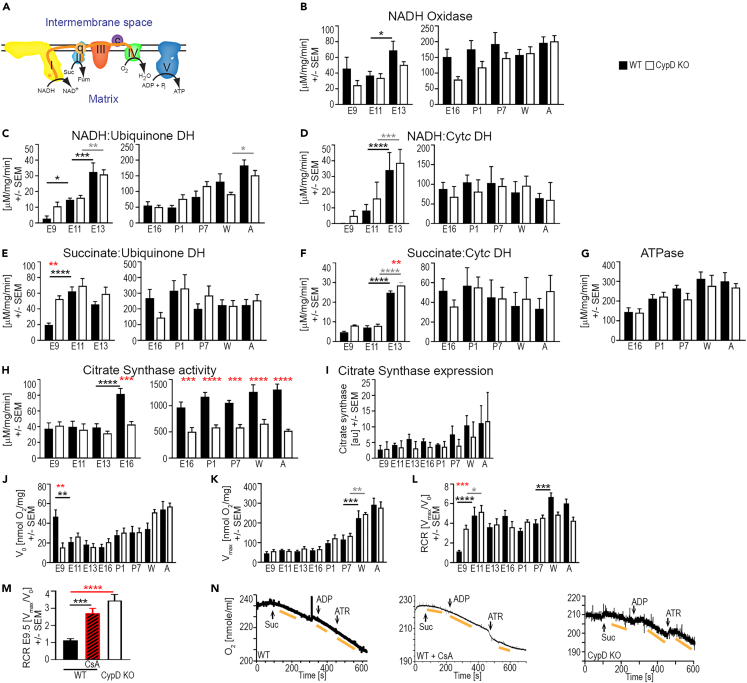
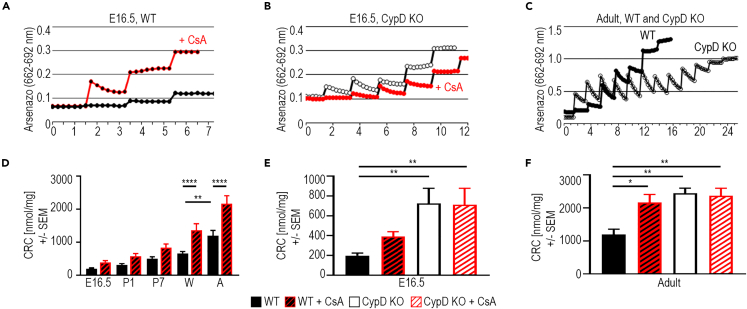
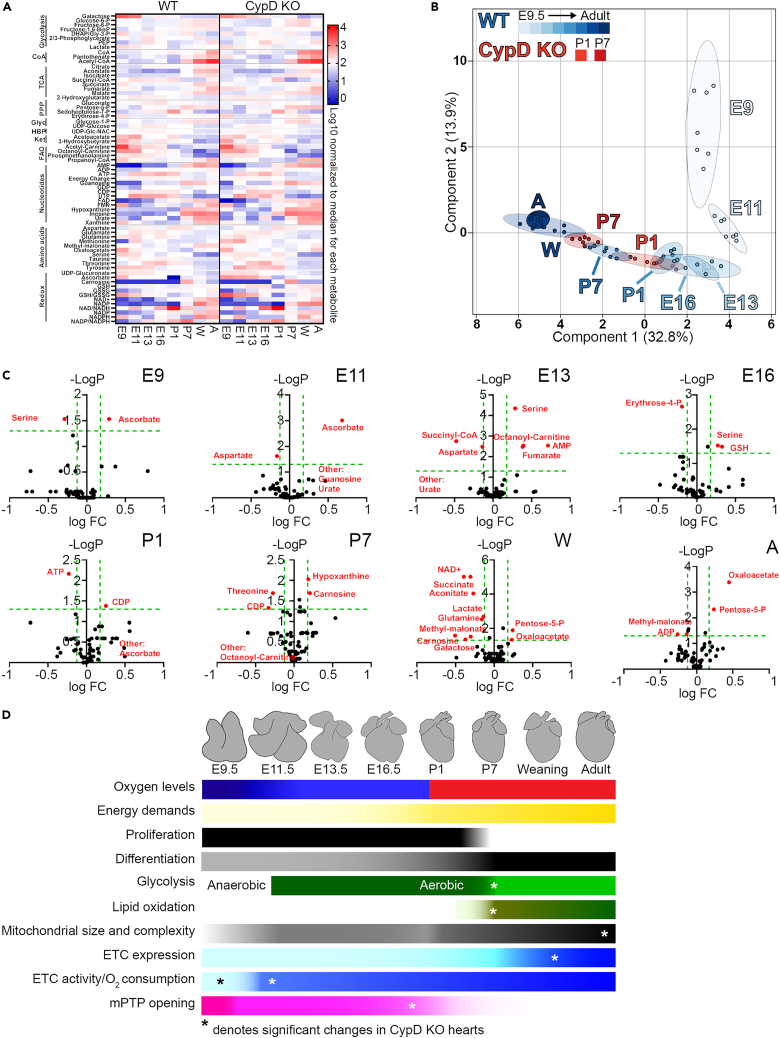
In the embryonic heart, the activation of the mitochondrial electron transport chain (ETC) coincides with the closure of the cyclophilin D (CypD) regulated mitochondrial permeability transition pore (mPTP). However, it remains to be established whether the absence of CypD has a regulatory effect on mitochondria during cardiac development. Using a variety of assays to analyze cardiac tissue from wildtype and CypD knockout mice from embryonic day (E)9.5 to adult, we found that mitochondrial structure, function, and metabolism show distinct transitions. Deletion of CypD altered the timing of these transitions as the mPTP was closed at all ages, leading to coupled ETC activity in the early embryo, decreased citrate synthase activity, and an altered metabolome particularly after birth. Our results suggest that manipulating CypD activity may control myocyte proliferation and differentiation and could be a tool to increase ATP production and cardiac function in immature hearts.
Keywords: Biological sciences; Cell biology; Metabolomics; Physiology.
© 2024 The Author(s).
Conflict of interest statement
GAPJr is the Inventor and the University of Rochester is the Assignee of United States Patent No. “US 10,179,161 B2” dated 1/15/19 and entitled “Compositions and methods for enhancing cardiac function in the neonate” that is related to the work reported herein. All other authors declare no competing interests.
Figures
Similar articles
-
Cyclophilin D Knock-Out Mice Show Enhanced Resistance to Osteoporosis and to Metabolic Changes Observed in Aging Bone.PLoS One. 2016 May 16;11(5):e0155709. doi: 10.1371/journal.pone.0155709. eCollection 2016. PLoS One. 2016. PMID: 27183225 Free PMC article.
-
Involvement of mitochondrial permeability transition pore (mPTP) in cardiac arrhythmias: Evidence from cyclophilin D knockout mice.Cell Calcium. 2016 Dec;60(6):363-372. doi: 10.1016/j.ceca.2016.09.001. Epub 2016 Sep 2. Cell Calcium. 2016. PMID: 27616659 Free PMC article.
-
Cyclophilin D-mediated Mitochondrial Permeability Transition Regulates Mitochondrial Function.Curr Pharm Des. 2023;29(8):620-629. doi: 10.2174/1381612829666230313111314. Curr Pharm Des. 2023. PMID: 36915987 Review.
-
Phosphorylation of cyclophilin D at serine 191 regulates mitochondrial permeability transition pore opening and cell death after ischemia-reperfusion.Cell Death Dis. 2020 Aug 19;11(8):661. doi: 10.1038/s41419-020-02864-5. Cell Death Dis. 2020. PMID: 32814770 Free PMC article.
-
Small-molecule inhibitors of cyclophilin D as potential therapeutics in mitochondria-related diseases.Med Res Rev. 2022 Sep;42(5):1822-1855. doi: 10.1002/med.21892. Epub 2022 May 16. Med Res Rev. 2022. PMID: 35575048 Review.
Cited by
-
Dysfunctional Electron Transport Chain Assembly in COXPD8.J Cardiovasc Dev Dis. 2025 Aug 20;12(8):318. doi: 10.3390/jcdd12080318. J Cardiovasc Dev Dis. 2025. PMID: 40863384 Free PMC article.
-
N-terminal cleavage of cyclophilin D boosts its ability to bind F-ATP synthase.Commun Biol. 2024 Nov 11;7(1):1486. doi: 10.1038/s42003-024-07172-8. Commun Biol. 2024. PMID: 39528709 Free PMC article.
-
The multifaceted role of mitochondria in cardiac function: insights and approaches.Cell Commun Signal. 2024 Oct 29;22(1):525. doi: 10.1186/s12964-024-01899-x. Cell Commun Signal. 2024. PMID: 39472951 Free PMC article. Review.
References
-
- Baker C.N., Ebert S.N. Development of aerobic metabolism in utero: Requirement for mitochondrial function during embryonic and fetal periods. OA Biotechnol. 2013;2:16.
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
