

Disruption of mitochondrial energy metabolism is a putative pathogenesis of Diamond-Blackfan anemia
- PMID: 38414864
- PMCID: PMC10897903
- DOI: 10.1016/j.isci.2024.109172
Disruption of mitochondrial energy metabolism is a putative pathogenesis of Diamond-Blackfan anemia
Abstract
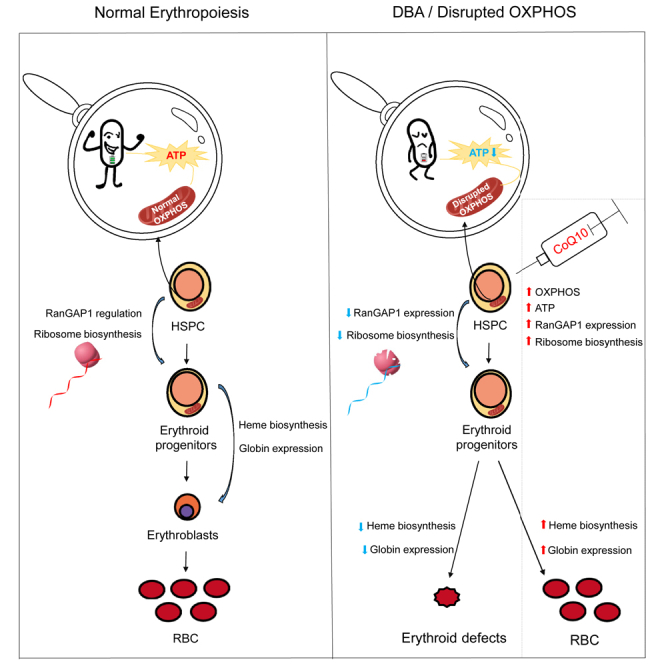
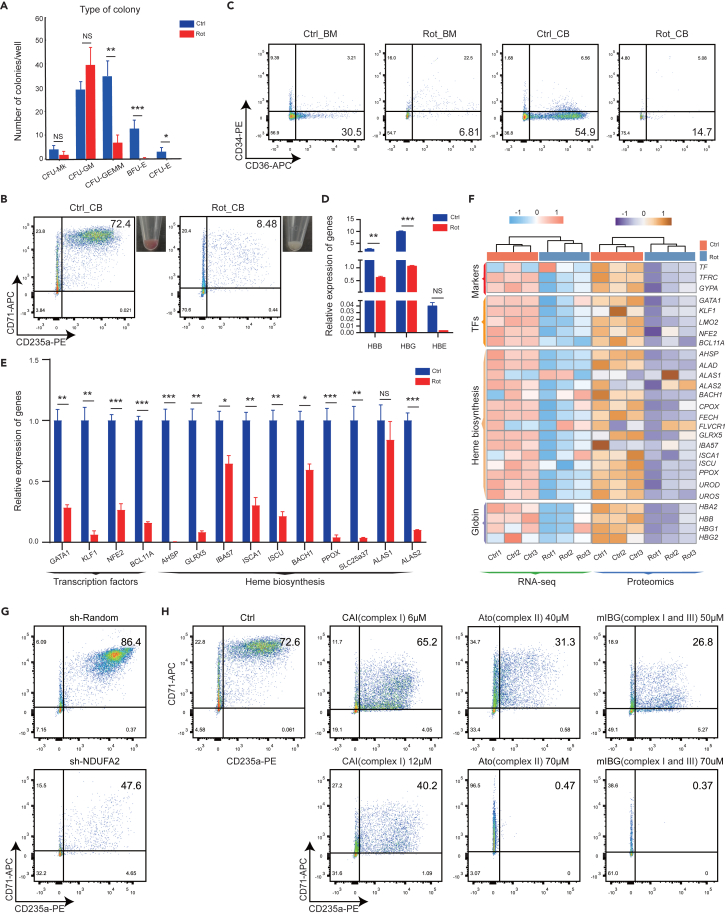
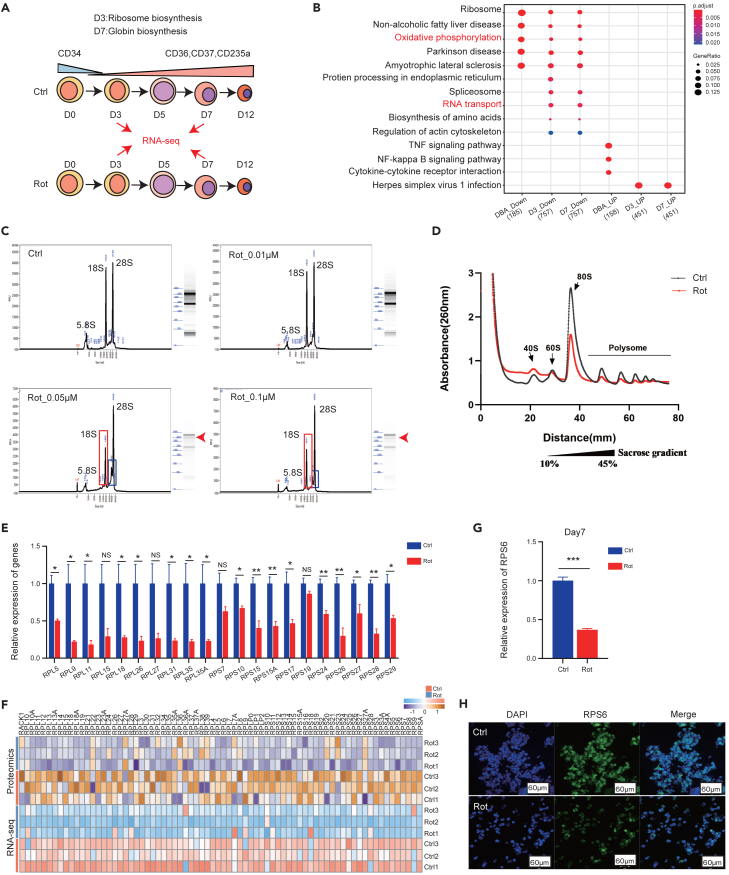
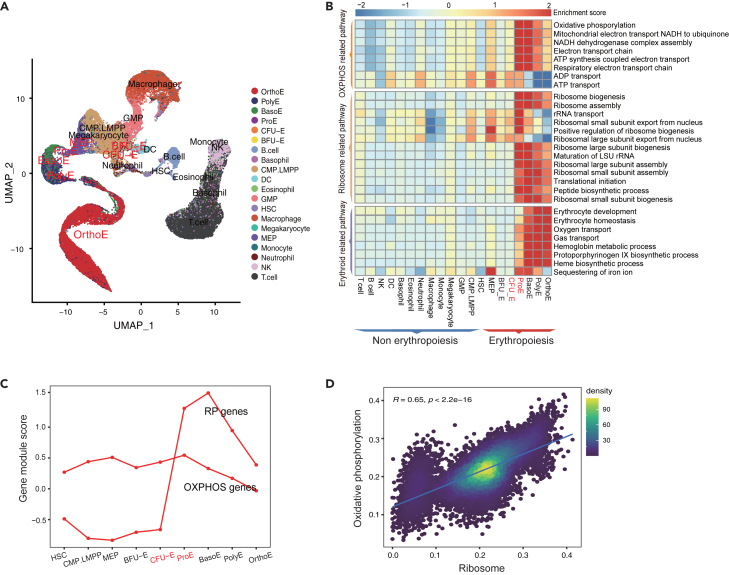
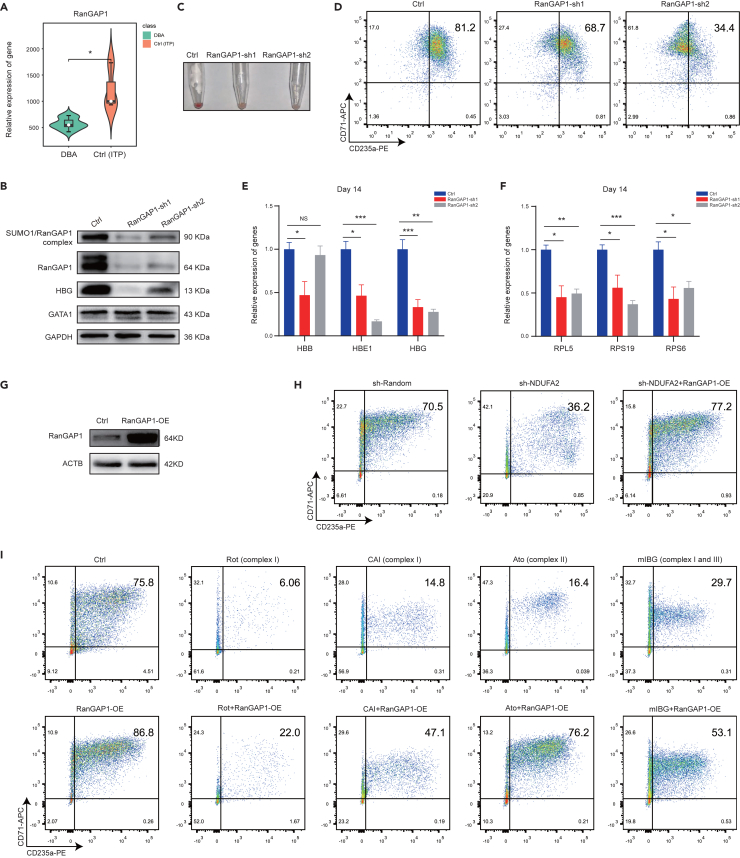
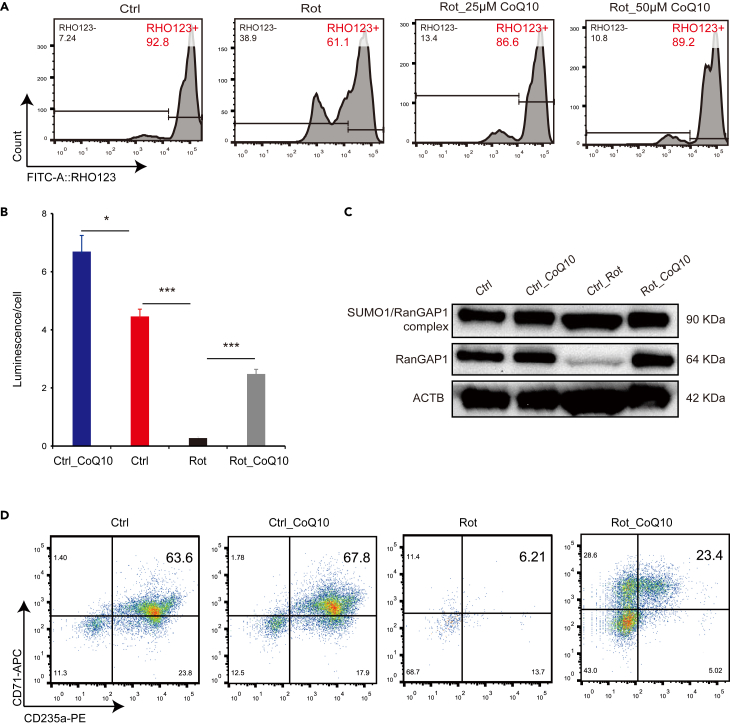
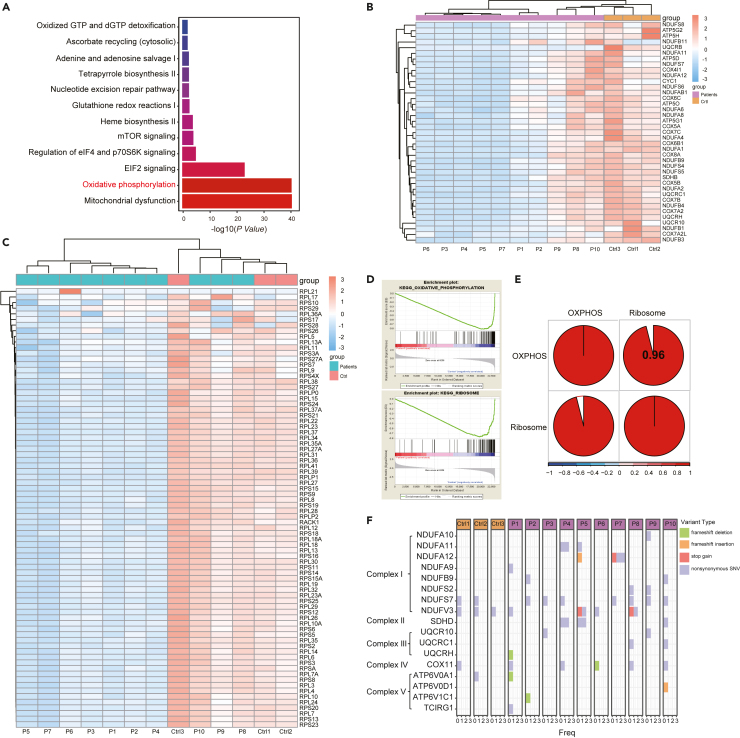
Energy metabolism in the context of erythropoiesis and related diseases remains largely unexplored. Here, we developed a primary cell model by differentiating hematopoietic stem progenitor cells toward the erythroid lineage and suppressing the mitochondrial oxidative phosphorylation (OXPHOS) pathway. OXPHOS suppression led to differentiation failure of erythroid progenitors and defects in ribosome biogenesis. Ran GTPase-activating protein 1 (RanGAP1) was identified as a target of mitochondrial OXPHOS for ribosomal defects during erythropoiesis. Overexpression of RanGAP1 largely alleviated erythroid defects resulting from OXPHOS suppression. Coenzyme Q10, an activator of OXPHOS, largely rescued erythroid defects and increased RanGAP1 expression. Patients with Diamond-Blackfan anemia (DBA) exhibited OXPHOS suppression and a concomitant suppression of ribosome biogenesis. RNA-seq analysis implied that the substantial mutation (approximately 10%) in OXPHOS genes accounts for OXPHOS suppression in these patients. Conclusively, OXPHOS disruption and the associated disruptive mitochondrial energy metabolism are linked to the pathogenesis of DBA.
Keywords: Cell biology; Cellular physiology; Developmental biology.
© 2024 The Author(s).
Conflict of interest statement
The authors declare that there are no competing financial interests in this study.
Figures
Similar articles
-
Enhanced alternative splicing of the FLVCR1 gene in Diamond Blackfan anemia disrupts FLVCR1 expression and function that are critical for erythropoiesis.Haematologica. 2008 Nov;93(11):1617-26. doi: 10.3324/haematol.13359. Epub 2008 Sep 24. Haematologica. 2008. PMID: 18815190
-
[Diamond-Blackfan anemia reveals the dark side of ribosome biogenesis].Med Sci (Paris). 2009 Jan;25(1):69-76. doi: 10.1051/medsci/200925169. Med Sci (Paris). 2009. PMID: 19154697 Review. French.
-
Ribosomal protein deficiency causes Tp53-independent erythropoiesis failure in zebrafish.Int J Biochem Cell Biol. 2014 Apr;49:1-7. doi: 10.1016/j.biocel.2014.01.006. Epub 2014 Jan 11. Int J Biochem Cell Biol. 2014. PMID: 24417973
-
Deficiency of ribosomal protein S26, which is mutated in a subset of patients with Diamond Blackfan anemia, impairs erythroid differentiation.Front Genet. 2022 Dec 12;13:1045236. doi: 10.3389/fgene.2022.1045236. eCollection 2022. Front Genet. 2022. PMID: 36579335 Free PMC article.
-
An update on the pathogenesis and diagnosis of Diamond-Blackfan anemia.F1000Res. 2018 Aug 29;7:F1000 Faculty Rev-1350. doi: 10.12688/f1000research.15542.1. eCollection 2018. F1000Res. 2018. PMID: 30228860 Free PMC article. Review.
Cited by
-
The involvement of mitochondria in erythrocyte pathology and diseases: from mechanisms to therapeutic strategies.Clin Exp Med. 2025 May 9;25(1):144. doi: 10.1007/s10238-024-01555-1. Clin Exp Med. 2025. PMID: 40343592 Free PMC article. Review.
References
-
- Chen M.L., Logan T.D., Hochberg M.L., Shelat S.G., Yu X., Wilding G.E., Tan W., Kujoth G.C., Prolla T.A., Selak M.A., et al. Erythroid dysplasia, megaloblastic anemia, and impaired lymphopoiesis arising from mitochondrial dysfunction. Blood. 2009;114:4045–4053. doi: 10.1182/blood-2008-08-169474. - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
