

Gamma oscillatory complexity conveys behavioral information in hippocampal networks
- PMID: 38418832
- PMCID: PMC10902292
- DOI: 10.1038/s41467-024-46012-5
Gamma oscillatory complexity conveys behavioral information in hippocampal networks
Abstract
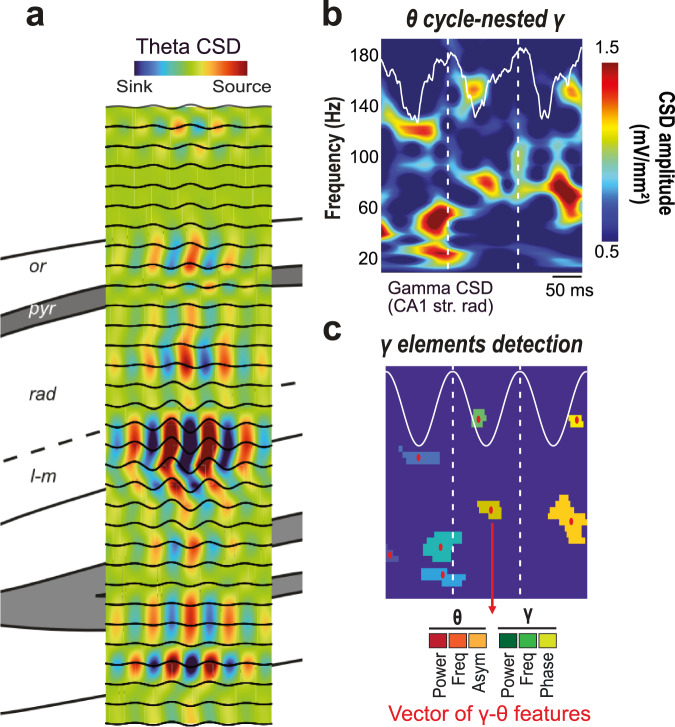
The hippocampus and entorhinal cortex exhibit rich oscillatory patterns critical for cognitive functions. In the hippocampal region CA1, specific gamma-frequency oscillations, timed at different phases of the ongoing theta rhythm, are hypothesized to facilitate the integration of information from varied sources and contribute to distinct cognitive processes. Here, we show that gamma elements -a multidimensional characterization of transient gamma oscillatory episodes- occur at any frequency or phase relative to the ongoing theta rhythm across all CA1 layers in male mice. Despite their low power and stochastic-like nature, individual gamma elements still carry behavior-related information and computational modeling suggests that they reflect neuronal firing. Our findings challenge the idea of rigid gamma sub-bands, showing that behavior shapes ensembles of irregular gamma elements that evolve with learning and depend on hippocampal layers. Widespread gamma diversity, beyond randomness, may thus reflect complexity, likely functional but invisible to classic average-based analyses.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures






References
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
