

Enhancing Human Treg Cell Induction through Engineered Dendritic Cells and Zinc Supplementation
- PMID: 38421704
- PMCID: PMC11015935
- DOI: 10.1615/CritRevImmunol.2023050325
Enhancing Human Treg Cell Induction through Engineered Dendritic Cells and Zinc Supplementation
Abstract
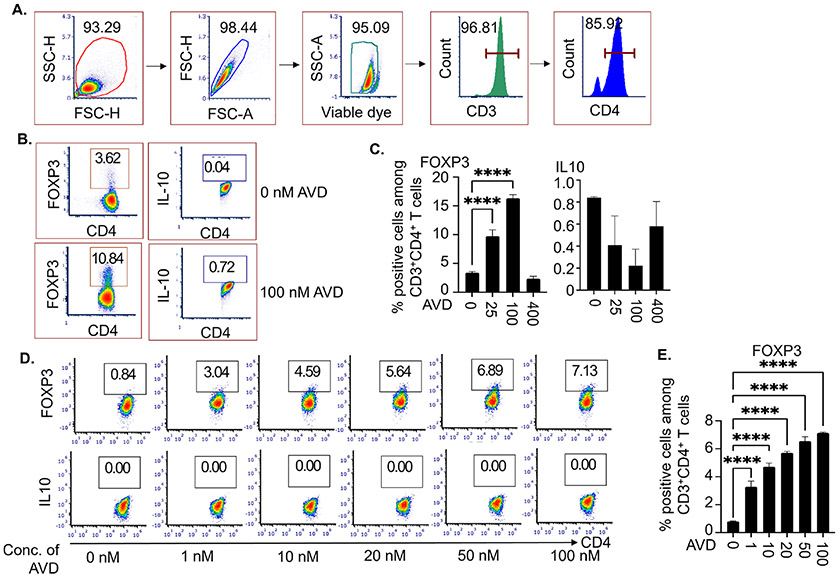
Regulatory T (Treg) cells hold promise for the ultimate cure of immune-mediated diseases. However, how to effectively restore Treg function in patients remains unknown. Previous reports suggest that activated dendritic cells (DCs) de novo synthesize locally high concentrations of 1,25-dihydroxy vitamin D, i.e., the active vitamin D or 1,25(OH)2D by upregulating the expression of 25-hydroxy vitamin D 1α-hydroxylase. Although 1,25(OH)2D has been shown to induce Treg cells, DC-derived 1,25(OH)2D only serves as a checkpoint to ensure well-balanced immune responses. Our animal studies have shown that 1,25(OH)2D requires high concentrations to generate Treg cells, which can cause severe side effects. In addition, our animal studies have also demonstrated that dendritic cells (DCs) overexpressing the 1α-hydroxylase de novo synthesize the effective Treg-inducing 1,25(OH)2D concentrations without causing the primary side effect of hypercalcemia (i.e., high blood calcium levels). This study furthers our previous animal studies and explores the efficacy of the la-hydroxylase-overexpressing DCs in inducing human CD4+FOXP3+regulatory T (Treg) cells. We discovered that the effective Treg-inducing doses of 1,25(OH)2D were within a range. Additionally, our data corroborated that the 1α-hydroxylase-overexpressing DCs synthesized 1,25(OH)2D within this concentration range in vivo, thus facilitating effective Treg cell induction. Moreover, this study demonstrated that 1α-hydroxylase expression levels were pivotal for DCs to induce Treg cells because physiological 25(OH)D levels were sufficient for the engineered but not parental DCs to enhance Treg cell induction. Interestingly, adding non-toxic zinc concentrations significantly augmented the Treg-inducing capacity of the engineered DCs. Our new findings offer a novel therapeutic avenue for immune-mediated human diseases, such as inflammatory bowel disease, type 1 diabetes, and multiple sclerosis, by integrating zinc with the 1α-hydroxylase-overexpressing DCs.
Figures






References
-
- Kim JM, Rasmussen JP, Rudensky AY. Regulatory T cells prevent catastrophic autoimmunity throughout the lifespan of mice. Nat Immunol. 2007;8(2):191–7.Epub 2006/12/01. - PubMed
-
- Fontenot JD, Gavin MA, Rudensky AY. Foxp3 programs the development and function of CD4+CD25+ regulatory T cells. Nat Immunol. 2003;4(4):330–6. Epub 2003/03/04. - PubMed
-
- Hori S, Nomura T, Sakaguchi S. Control of regulatory T cell development by the transcription factor Foxp3. Science. 2003;299(5609):1057–61. Epub 2003/01/11. - PubMed
-
- Bennett CL, Christie J, Ramsdell F, Brunkow ME, Ferguson PJ, Whitesell L, Kelly TE, Saulsbury FT, Chance PF, Ochs HD. The immune dysregulation, polyendocrinopathy, enteropathy, X-linked syndrome (IPEX) is caused by mutations of FOXP3. Nat Genet. 2001;27(1):20–1. Epub 2001/01/04. - PubMed
-
- Khattri R, Cox T, Yasayko SA, Ramsdell F. An essential role for Scurfin in CD4+CD25+ T regulatory cells. Nat Immunol. 2003;4(4):337–42. Epub 2003/03/04. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials