

Synergistic toxicity with copper contributes to NAT2-associated isoniazid toxicity
- PMID: 38424191
- PMCID: PMC10984958
- DOI: 10.1038/s12276-024-01172-8
Synergistic toxicity with copper contributes to NAT2-associated isoniazid toxicity
Abstract
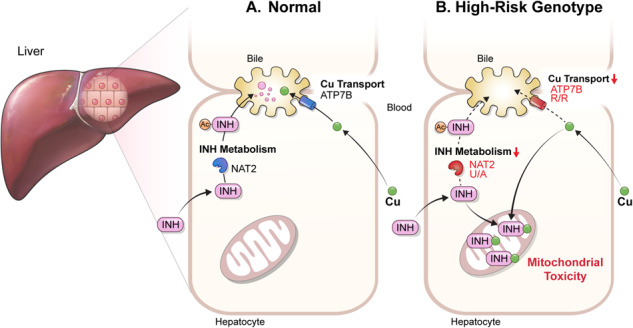
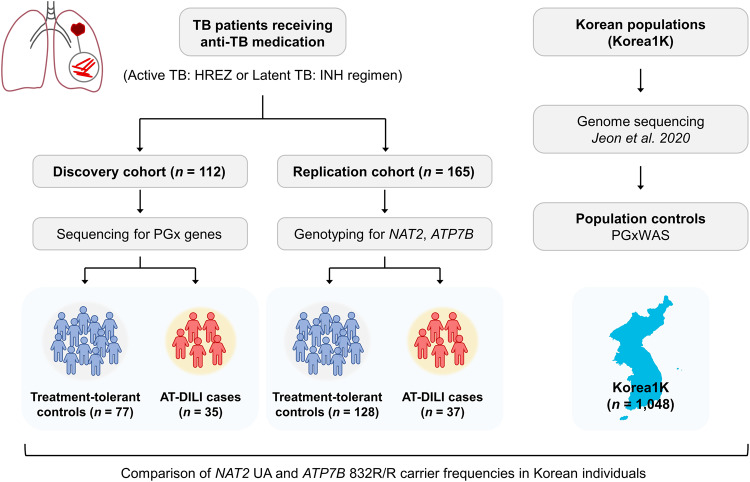
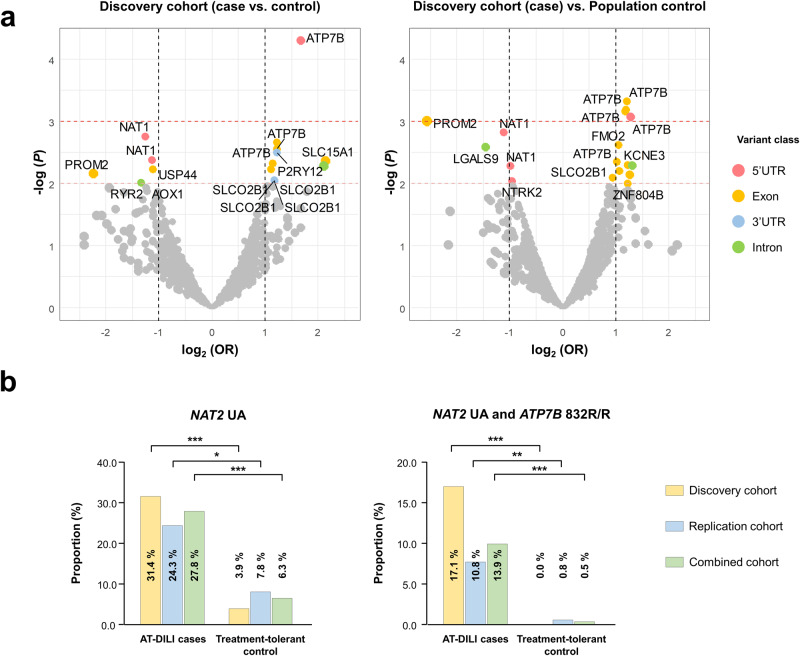
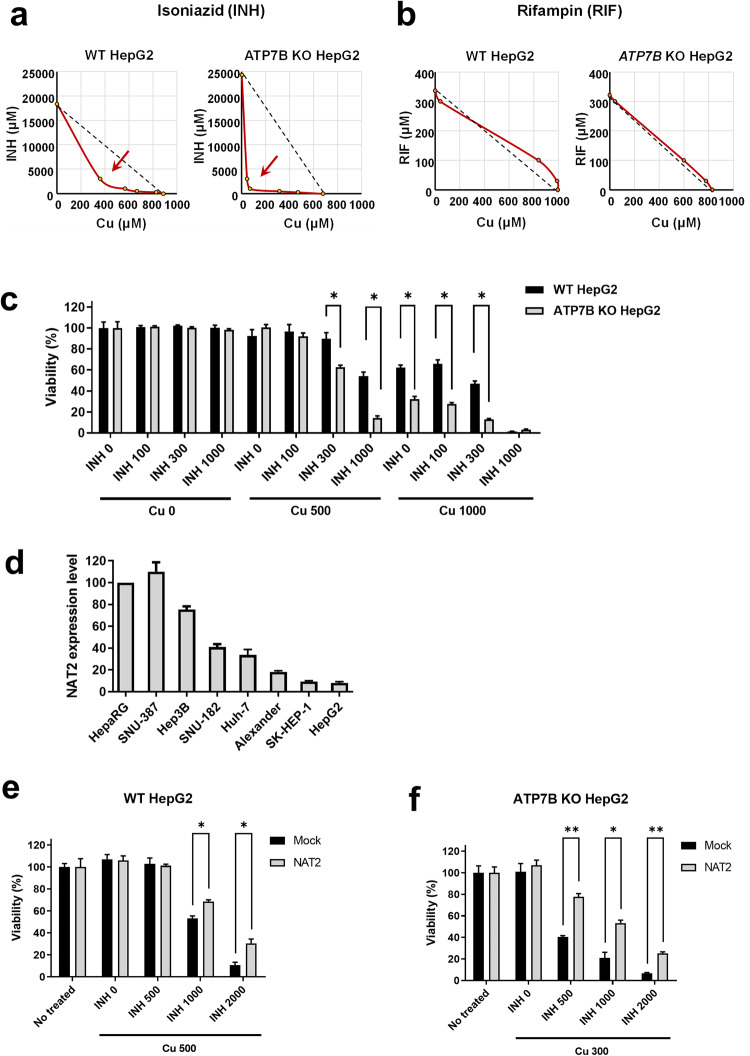
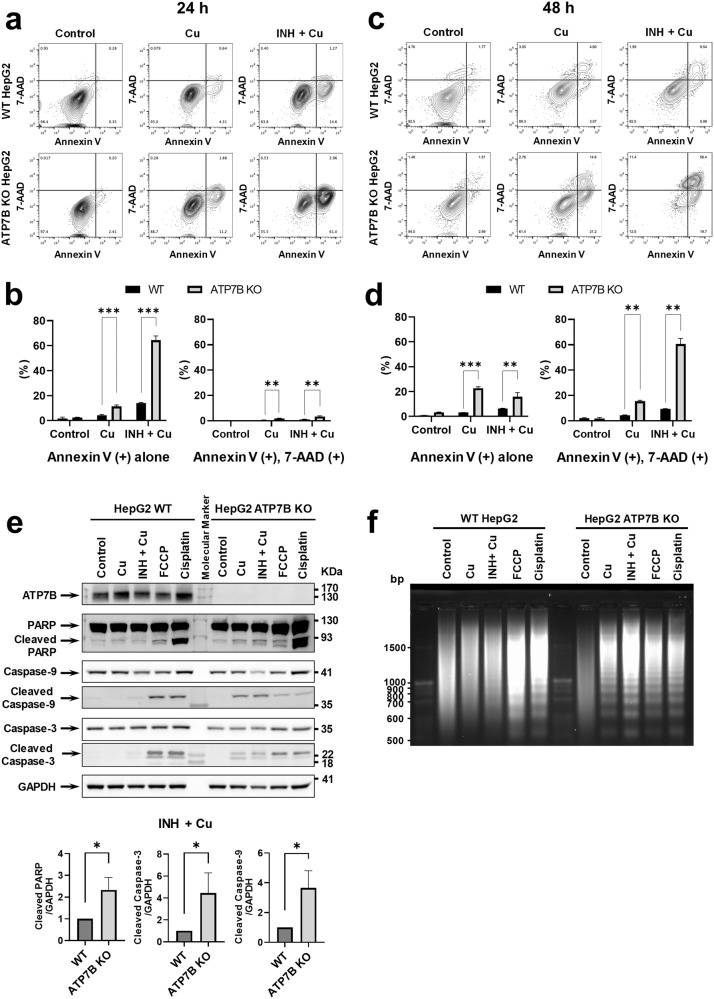
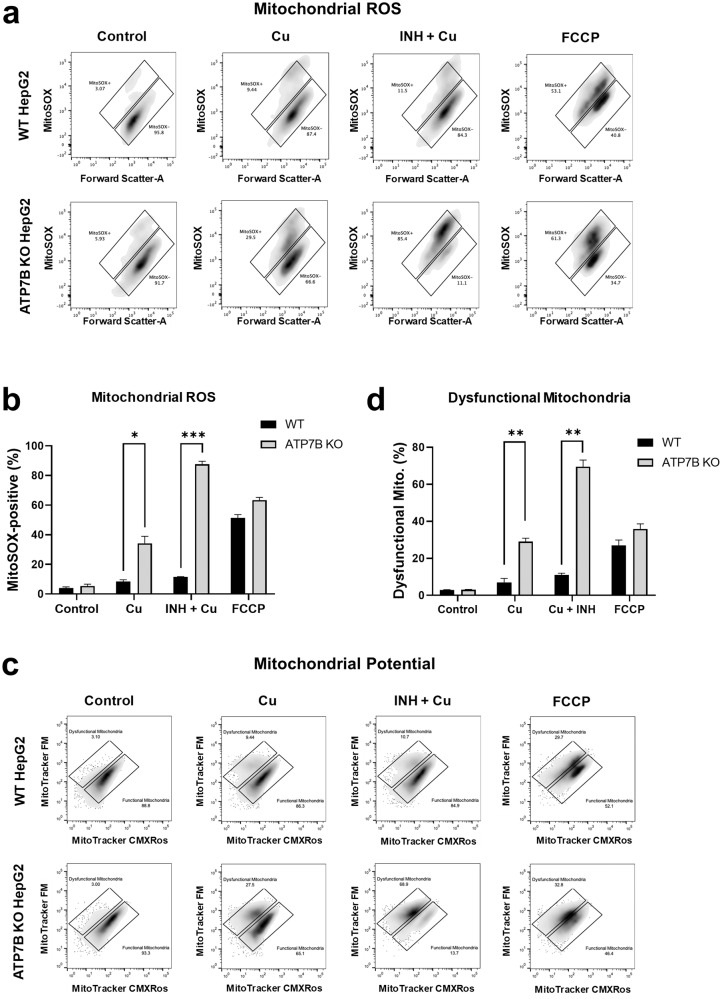
Anti-tuberculosis (AT) medications, including isoniazid (INH), can cause drug-induced liver injury (DILI), but the underlying mechanism remains unclear. In this study, we aimed to identify genetic factors that may increase the susceptibility of individuals to AT-DILI and to examine genetic interactions that may lead to isoniazid (INH)-induced hepatotoxicity. We performed a targeted sequencing analysis of 380 pharmacogenes in a discovery cohort of 112 patients (35 AT-DILI patients and 77 controls) receiving AT treatment for active tuberculosis. Pharmacogenome-wide association analysis was also conducted using 1048 population controls (Korea1K). NAT2 and ATP7B genotypes were analyzed in a replication cohort of 165 patients (37 AT-DILI patients and 128 controls) to validate the effects of both risk genotypes. NAT2 ultraslow acetylators (UAs) were found to have a greater risk of AT-DILI than other genotypes (odds ratio [OR] 5.6 [95% confidence interval; 2.5-13.2], P = 7.2 × 10-6). The presence of ATP7B gene 832R/R homozygosity (rs1061472) was found to co-occur with NAT2 UA in AT-DILI patients (P = 0.017) and to amplify the risk in NAT2 UA (OR 32.5 [4.5-1423], P = 7.5 × 10-6). In vitro experiments using human liver-derived cell lines (HepG2 and SNU387 cells) revealed toxic synergism between INH and Cu, which were strongly augmented in cells with defective NAT2 and ATP7B activity, leading to increased mitochondrial reactive oxygen species generation, mitochondrial dysfunction, DNA damage, and apoptosis. These findings link the co-occurrence of ATP7B and NAT2 genotypes to the risk of INH-induced hepatotoxicity, providing novel mechanistic insight into individual AT-DILI susceptibility. Yoon et al. showed that individuals who carry NAT2 UAs and ATP7B 832R/R genotypes are at increased risk of developing isoniazid hepatotoxicity, primarily due to the increased synergistic toxicity between isoniazid and copper, which exacerbates mitochondrial dysfunction-related apoptosis.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
