

Targeted deletion of CD244 on monocytes promotes differentiation into anti-tumorigenic macrophages and potentiates PD-L1 blockade in melanoma
- PMID: 38424542
- PMCID: PMC10903025
- DOI: 10.1186/s12943-024-01936-w
Targeted deletion of CD244 on monocytes promotes differentiation into anti-tumorigenic macrophages and potentiates PD-L1 blockade in melanoma
Abstract
Background: In the myeloid compartment of the tumor microenvironment, CD244 signaling has been implicated in immunosuppressive phenotype of monocytes. However, the precise molecular mechanism and contribution of CD244 to tumor immunity in monocytes/macrophages remains elusive due to the co-existing lymphoid cells expressing CD244.
Methods: To directly assess the role of CD244 in tumor-associated macrophages, monocyte-lineage-specific CD244-deficient mice were generated using cre-lox recombination and challenged with B16F10 melanoma. The phenotype and function of tumor-infiltrating macrophages along with antigen-specific CD8 T cells were analyzed by flow cytometry and single cell RNA sequencing data analysis, and the molecular mechanism underlying anti-tumorigenic macrophage differentiation, antigen presentation, phagocytosis was investigated ex vivo. Finally, the clinical feasibility of CD244-negative monocytes as a therapeutic modality in melanoma was confirmed by adoptive transfer experiments.
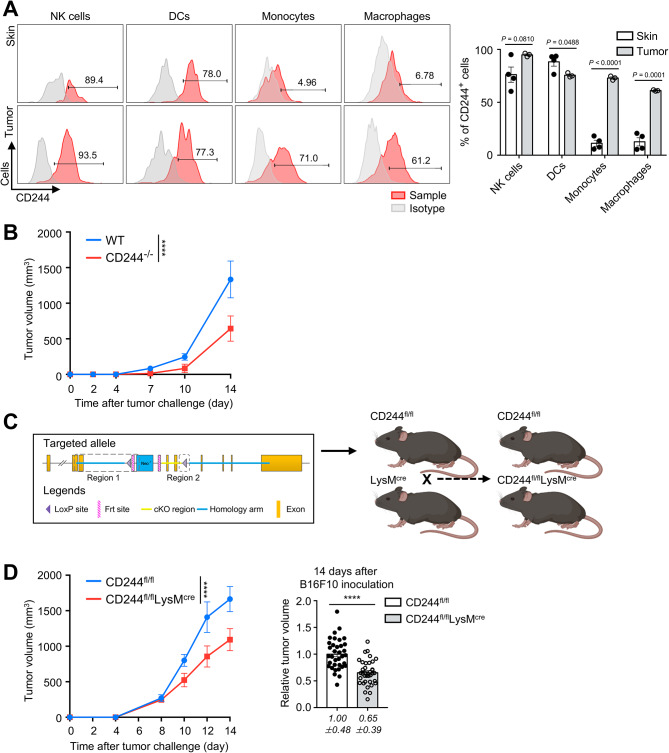
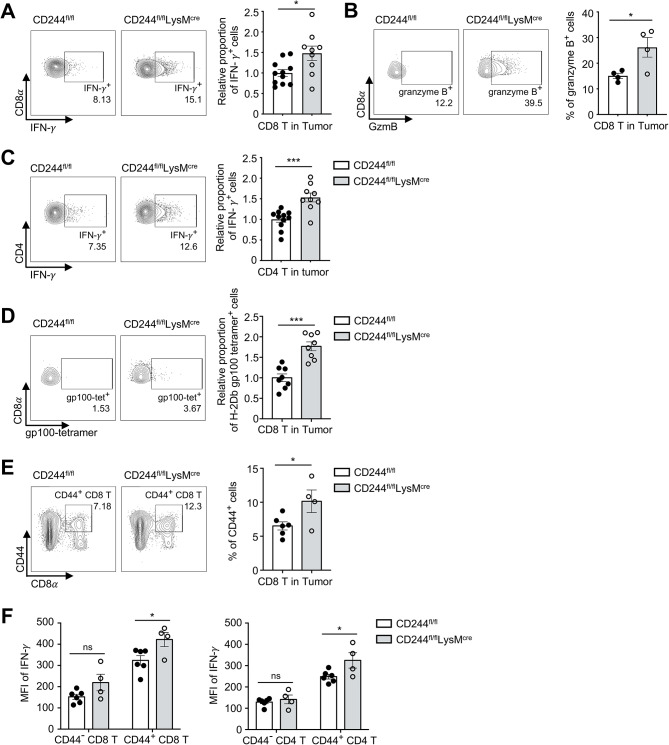
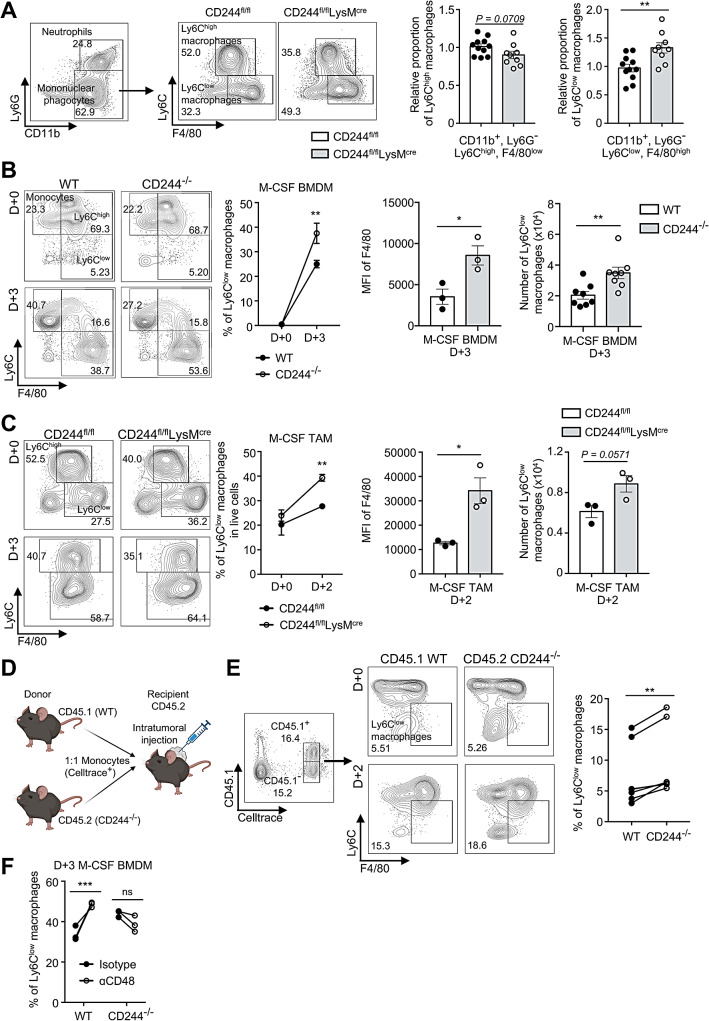
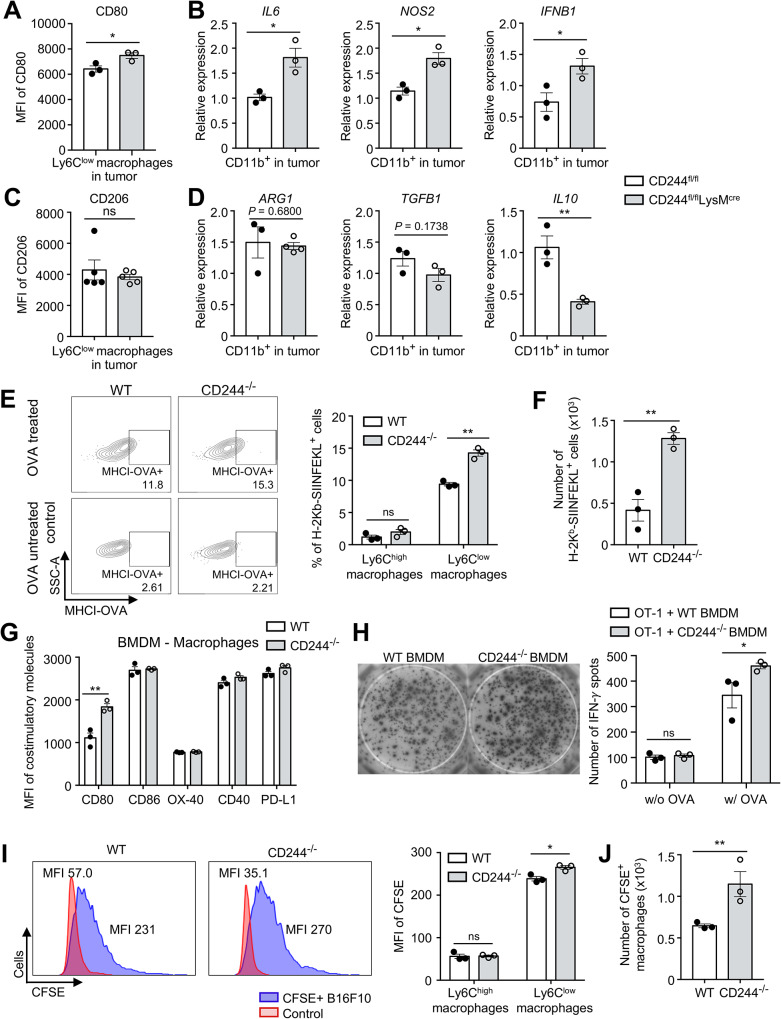
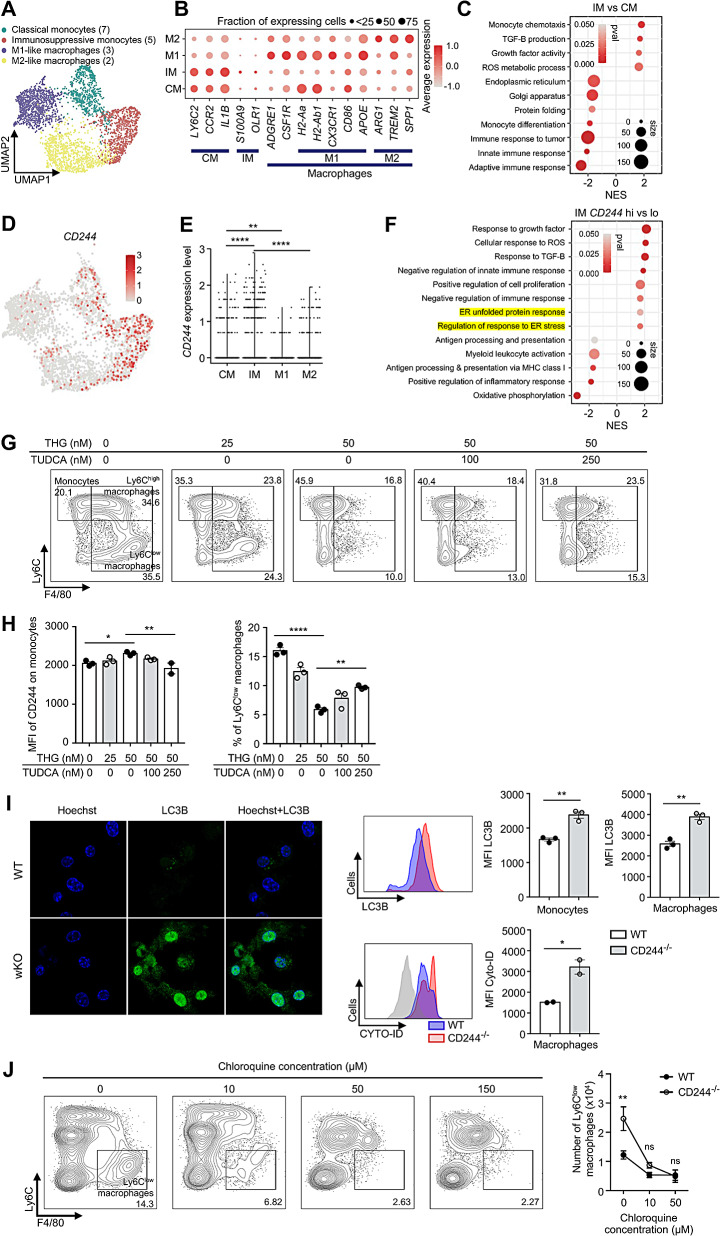
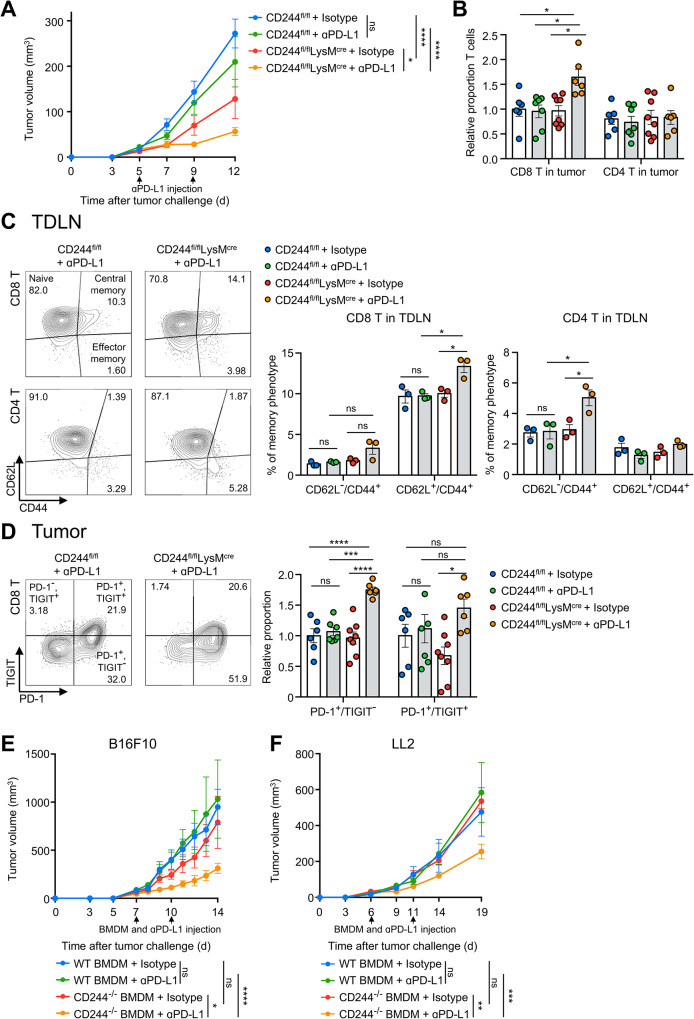
Results: CD244fl/flLysMcre mice demonstrated a significant reduction in tumor volume (61% relative to that of the CD244fl/fl control group) 14 days after tumor implantation. Within tumor mass, CD244fl/flLysMcre mice also showed higher percentages of Ly6Clow macrophages, along with elevated gp100+IFN-γ+ CD8 T cells. Flow cytometry and RNA sequencing data demonstrated that ER stress resulted in increased CD244 expression on monocytes. This, in turn, impeded the generation of anti-tumorigenic Ly6Clow macrophages, phagocytosis and MHC-I antigen presentation by suppressing autophagy pathways. Combining anti-PD-L1 antibody with CD244-/- bone marrow-derived macrophages markedly improved tumor rejection compared to the anti-PD-L1 antibody alone or in combination with wild-type macrophages. Consistent with the murine data, transcriptome analysis of human melanoma tissue single-cell RNA-sequencing dataset revealed close association between CD244 and the inhibition of macrophage maturation and function. Furthermore, the presence of CD244-negative monocytes/macrophages significantly increased patient survival in primary and metastatic tumors.
Conclusion: Our study highlights the novel role of CD244 on monocytes/macrophages in restraining anti-tumorigenic macrophage generation and tumor antigen-specific T cell response in melanoma. Importantly, our findings suggest that CD244-deficient macrophages could potentially be used as a therapeutic agent in combination with immune checkpoint inhibitors. Furthermore, CD244 expression in monocyte-lineage cells serve as a prognostic marker in cancer patients.
Keywords: CD244; Differentiation; Immune checkpoint blockade; Macrophage-based cell therapy; Macrophages; Melanoma.
© 2024. The Author(s).
Conflict of interest statement
JSK and KML are named inventors on a patent for a pharmaceutical composition to prevent or treat cancer-comprising monocytes or macrophages that inhibit CD244 expression or activity as an active ingredient (KR10-2022-0067313, PCT/KR2023/007574).
Figures

References
-
- Lee KM, Chuang E, Griffin M, Khattri R, Hong DK, Zhang WG, et al. Molecular basis of T cell inactivation by CTLA-4. Science. 1998;282(5397):2263–6. - PubMed
-
- Cai GF, Freeman GJ. The CD160, BTLA, LIGHT/HVEM pathway: a bidirectional switch regulating T-cell activation. Immunol Rev. 2009;229:244–58. - PubMed
-
- Calabro L, Danielli R, Sigalotti L, Maio M. Clinical studies with Anti-CTLA-4 antibodies in non-melanoma indications. Semin Oncol. 2010;37(5):460–7. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
