

PIEZO1 is a distal nephron mechanosensor and is required for flow-induced K+ secretion
- PMID: 38426496
- PMCID: PMC10904061
- DOI: 10.1172/JCI174806
PIEZO1 is a distal nephron mechanosensor and is required for flow-induced K+ secretion
Abstract
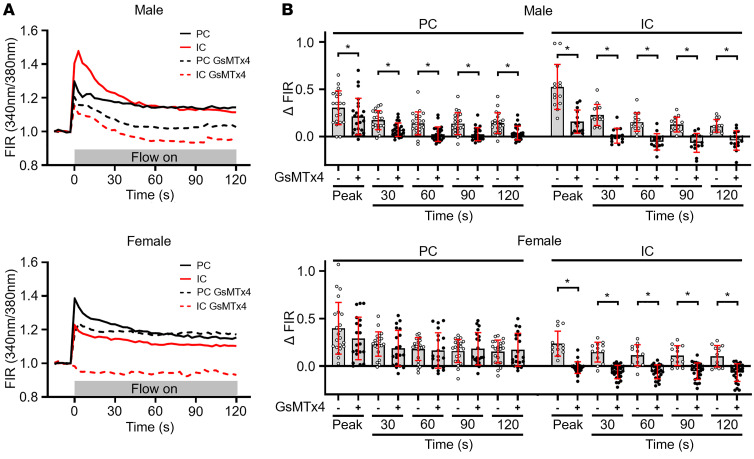
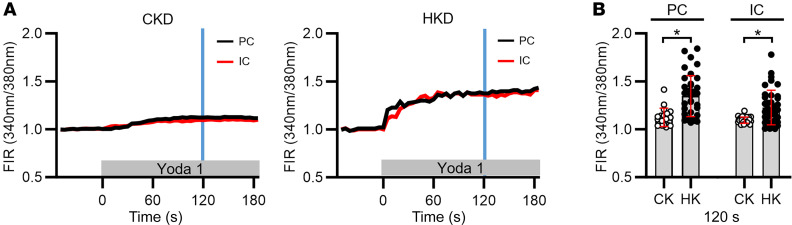
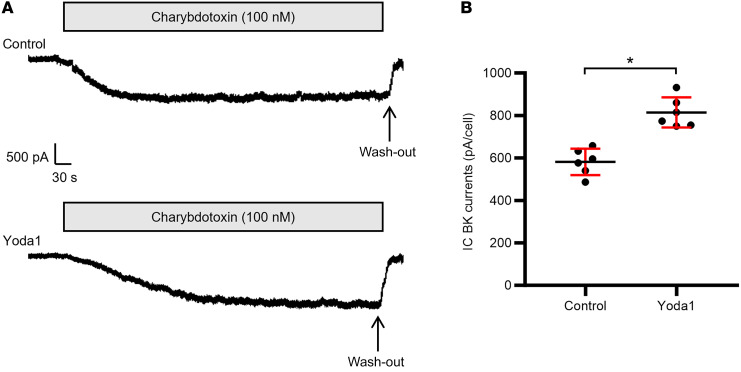
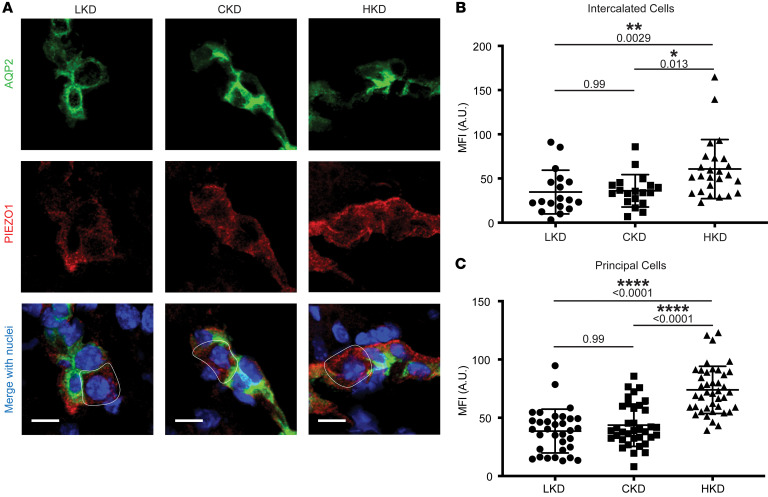
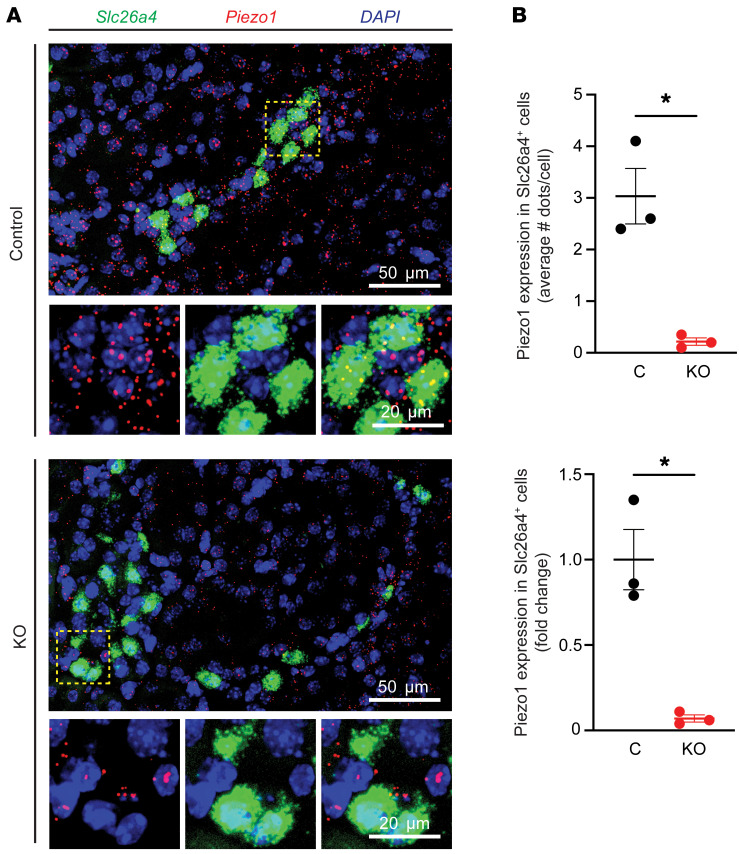
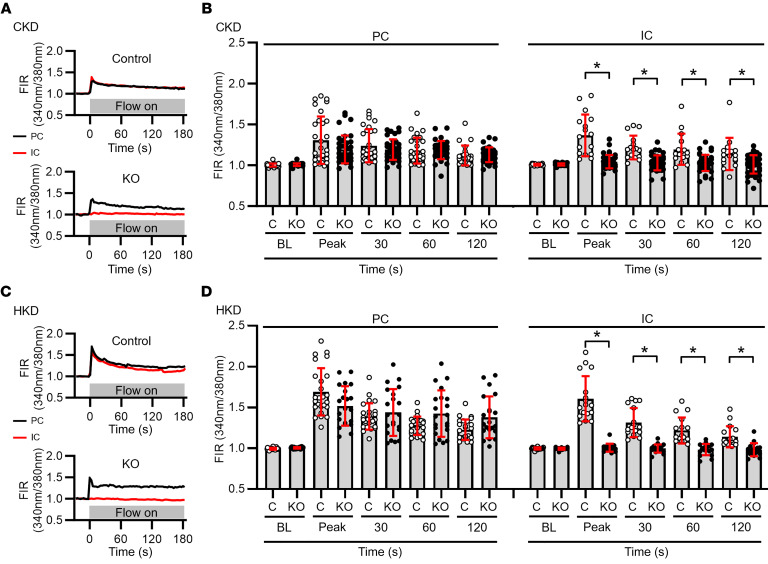
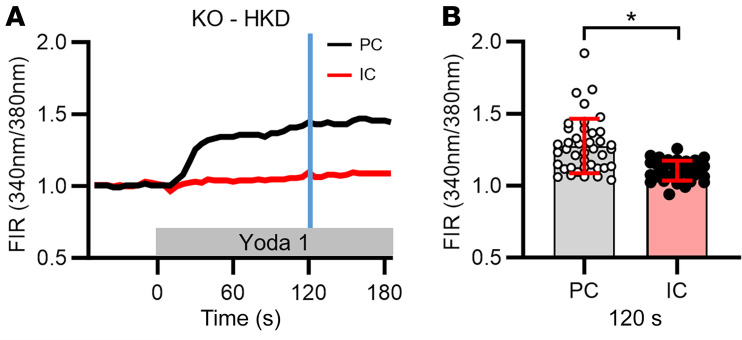
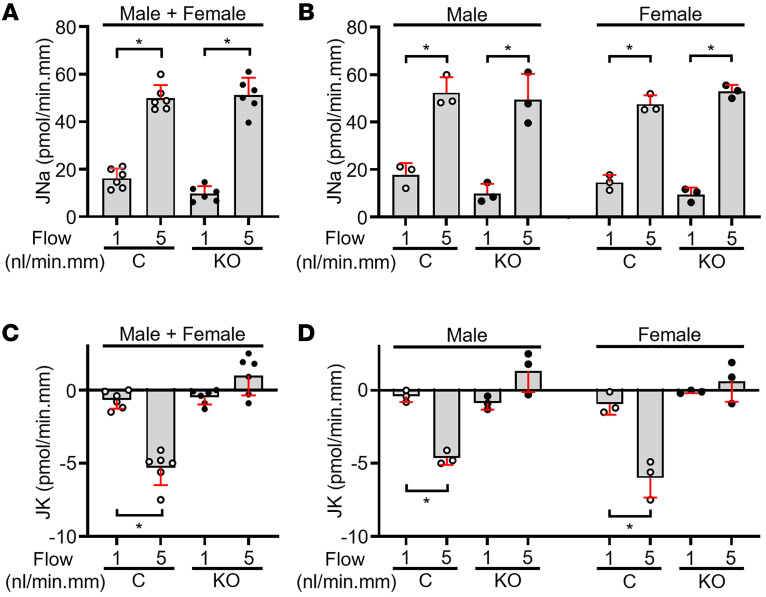
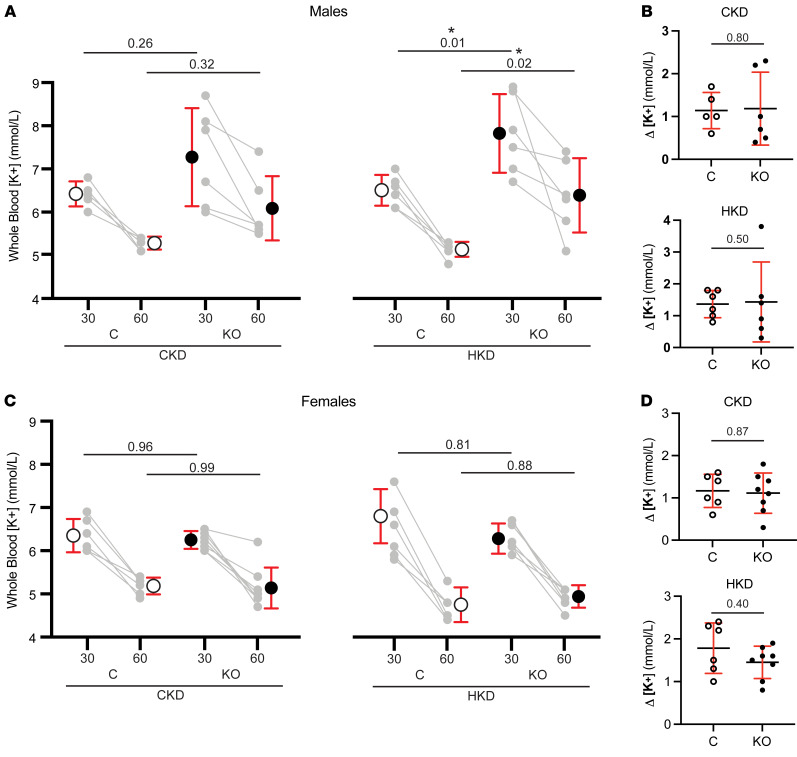
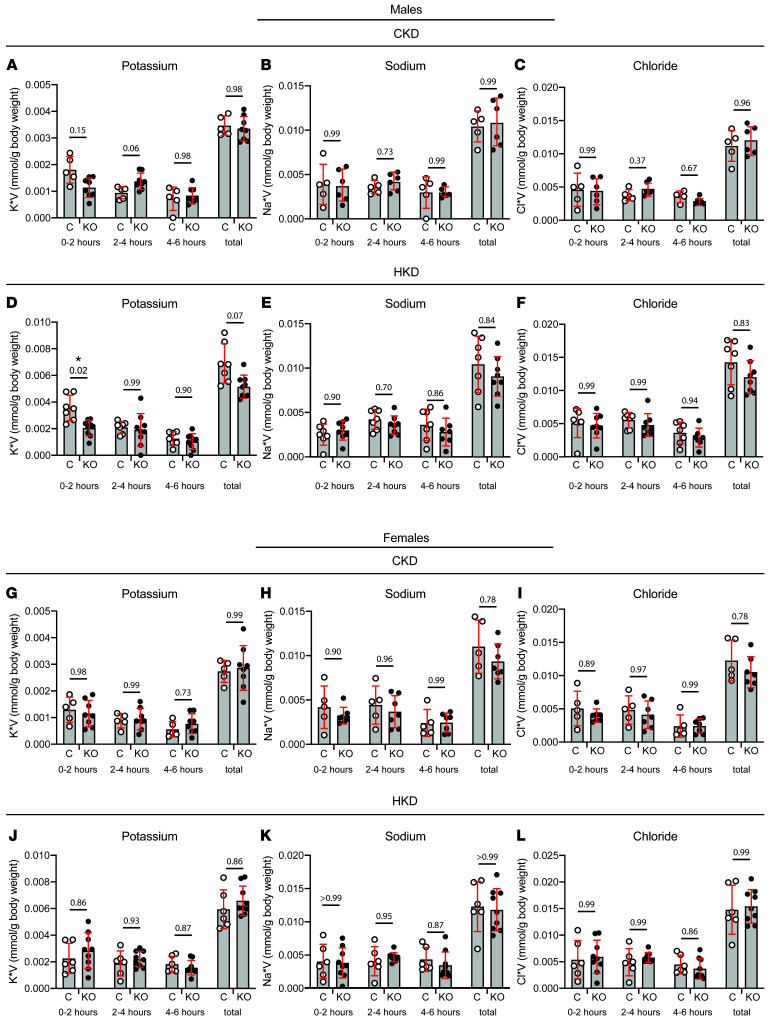
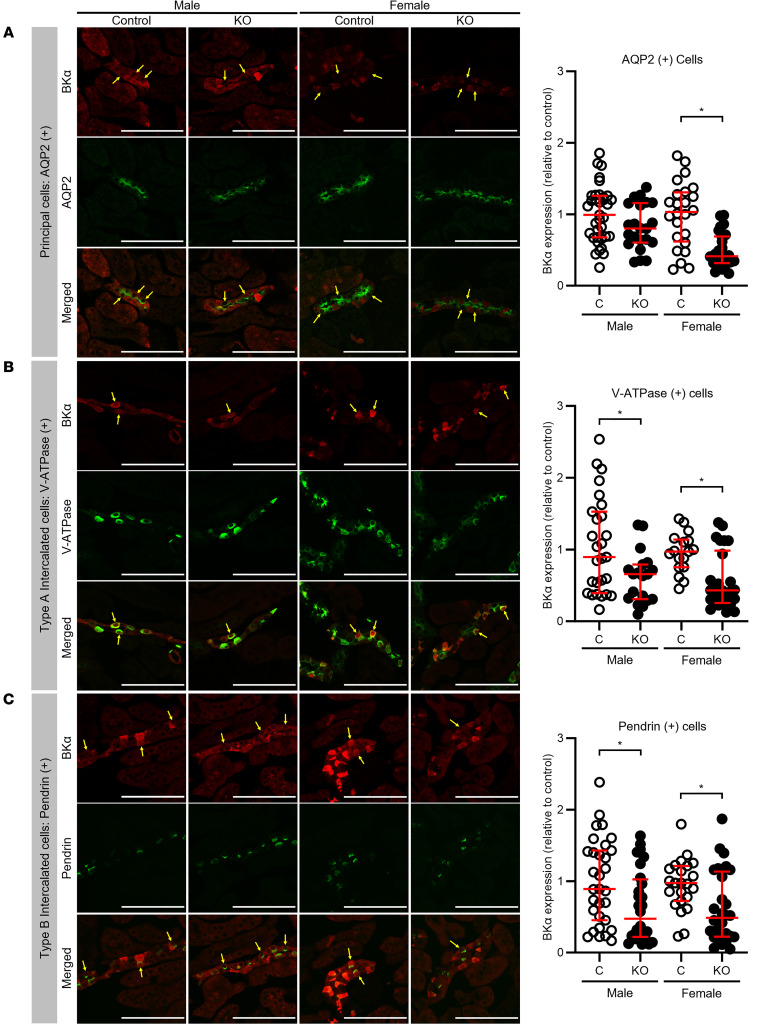
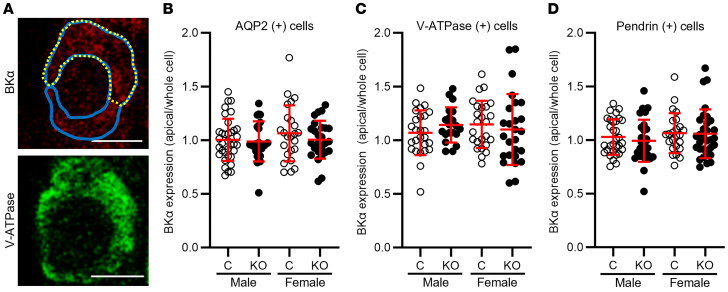
Ca2+-activated BK channels in renal intercalated cells (ICs) mediate luminal flow-induced K+ secretion (FIKS), but how ICs sense increased flow remains uncertain. We examined whether PIEZO1, a mechanosensitive Ca2+-permeable channel expressed in the basolateral membranes of ICs, is required for FIKS. In isolated cortical collecting ducts (CCDs), the mechanosensitive cation-selective channel inhibitor GsMTx4 dampened flow-induced increases in intracellular Ca2+ concentration ([Ca2+]i), whereas the PIEZO1 activator Yoda1 increased [Ca2+]i and BK channel activity. CCDs from mice fed a high-K+ (HK) diet exhibited a greater Yoda1-dependent increase in [Ca2+]i than CCDs from mice fed a control K+ diet. ICs in CCDs isolated from mice with a targeted gene deletion of Piezo1 in ICs (IC-Piezo1-KO) exhibited a blunted [Ca2+]i response to Yoda1 or increased flow, with an associated loss of FIKS in CCDs. Male IC-Piezo1-KO mice selectively exhibited an increased blood [K+] in response to an oral K+ bolus and blunted urinary K+ excretion following a volume challenge. Whole-cell expression of BKα subunit was reduced in ICs of IC-Piezo1-KO mice fed an HK diet. We conclude that PIEZO1 mediates flow-induced basolateral Ca2+ entry into ICs, is upregulated in the CCD in response to an HK diet, and is necessary for FIKS.
Keywords: Calcium channels; Cell biology; Epithelial transport of ions and water; Nephrology; Potassium channels.
Figures
References
-
- Palmer LG, Frindt G. Regulation of apical K channels in rat cortical collecting tubule during changes in dietary K intake. Am J Physiol. 1999;277(5):F805–F812. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
