

Cancer neoepitopes viewed through negative selection and peripheral tolerance: a new path to cancer vaccines
- PMID: 38426497
- PMCID: PMC10904052
- DOI: 10.1172/JCI176740
Cancer neoepitopes viewed through negative selection and peripheral tolerance: a new path to cancer vaccines
Abstract
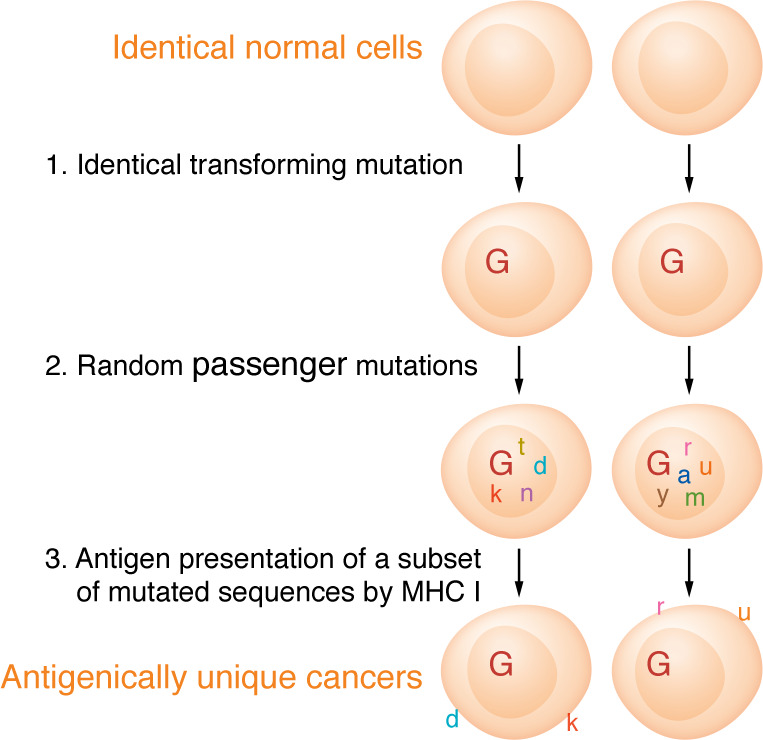
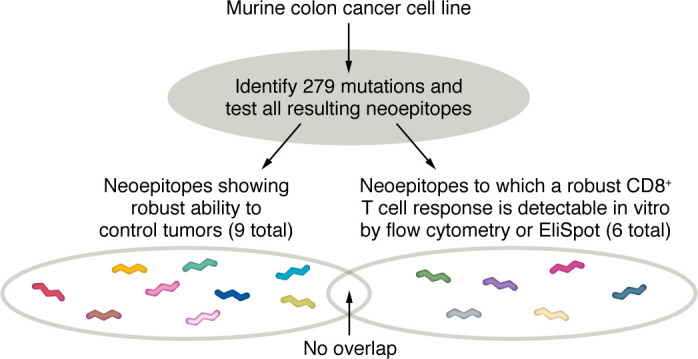
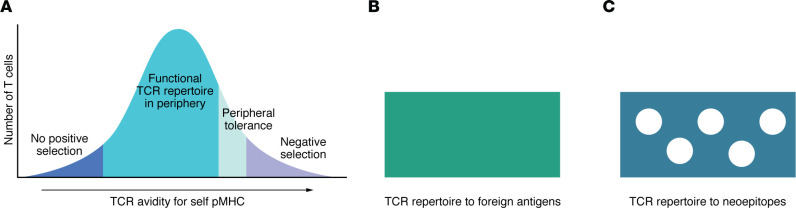
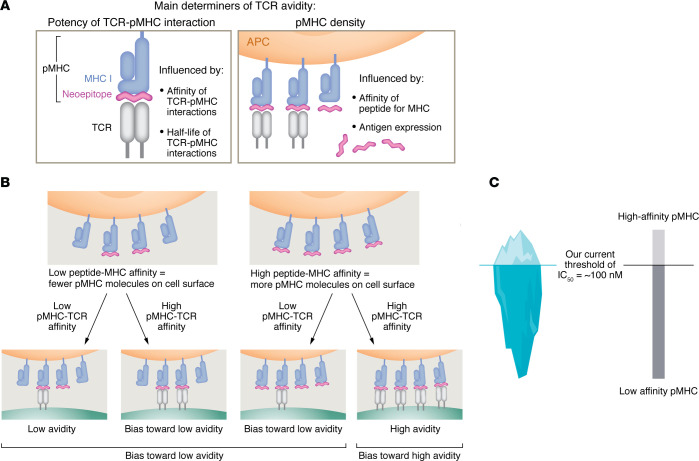
A proportion of somatic mutations in tumors create neoepitopes that can prime T cell responses that target the MHC I-neoepitope complexes on tumor cells, mediating tumor control or rejection. Despite the compelling centrality of neoepitopes to cancer immunity, we know remarkably little about what constitutes a neoepitope that can mediate tumor control in vivo and what distinguishes such a neoepitope from the vast majority of similar candidate neoepitopes that are inefficacious in vivo. Studies in mice as well as clinical trials have begun to reveal the unexpected paradoxes in this area. Because cancer neoepitopes straddle that ambiguous ground between self and non-self, some rules that are fundamental to immunology of frankly non-self antigens, such as viral or model antigens, do not appear to apply to neoepitopes. Because neoepitopes are so similar to self-epitopes, with only small changes that render them non-self, immune response to them is regulated at least partially the way immune response to self is regulated. Therefore, neoepitopes are viewed and understood here through the clarifying lens of negative thymic selection. Here, the emergent questions in the biology and clinical applications of neoepitopes are discussed critically and a mechanistic and testable framework that explains the complexity and translational potential of these wonderful antigens is proposed.
Conflict of interest statement
Figures
Similar articles
-
Population-level distribution and putative immunogenicity of cancer neoepitopes.BMC Cancer. 2018 Apr 13;18(1):414. doi: 10.1186/s12885-018-4325-6. BMC Cancer. 2018. PMID: 29653567 Free PMC article.
-
Prediction of cancer neoepitopes needs new rules.Semin Immunol. 2020 Feb;47:101387. doi: 10.1016/j.smim.2020.101387. Epub 2020 Jan 14. Semin Immunol. 2020. PMID: 31952902 Review.
-
Mass spectrometry driven exploration reveals nuances of neoepitope-driven tumor rejection.JCI Insight. 2019 Jun 20;5(14):e129152. doi: 10.1172/jci.insight.129152. JCI Insight. 2019. PMID: 31219806 Free PMC article.
-
Structural and physical features that distinguish tumor-controlling from inactive cancer neoepitopes.Proc Natl Acad Sci U S A. 2023 Dec 19;120(51):e2312057120. doi: 10.1073/pnas.2312057120. Epub 2023 Dec 12. Proc Natl Acad Sci U S A. 2023. PMID: 38085776 Free PMC article.
-
Targeting the Heterogeneity of Cancer with Individualized Neoepitope Vaccines.Clin Cancer Res. 2016 Apr 15;22(8):1885-96. doi: 10.1158/1078-0432.CCR-15-1509. Clin Cancer Res. 2016. PMID: 27084742 Review.
Cited by
-
Hypothesis: the generation of T cells directed against neoepitopes employing immune-mediating agents other than neoepitope vaccines.J Immunother Cancer. 2024 Jul 8;12(7):e009595. doi: 10.1136/jitc-2024-009595. J Immunother Cancer. 2024. PMID: 38977329 Free PMC article.
-
A frameshift-generated cancer neoepitope that controls tumor burden in prophylaxis as well as therapy.J Immunol. 2025 Jun 1;214(6):1123-1132. doi: 10.1093/jimmun/vkaf016. J Immunol. 2025. PMID: 40209093 Free PMC article.
-
Vaccines in cancer treatment and prevention: the time is now.J Clin Invest. 2025 Jul 1;135(13):e195673. doi: 10.1172/JCI195673. eCollection 2025 Jul 1. J Clin Invest. 2025. PMID: 40590227 Free PMC article. No abstract available.
-
Dynamic allostery in the peptide/MHC complex enables TCR neoantigen selectivity.Nat Commun. 2025 Jan 20;16(1):849. doi: 10.1038/s41467-025-56004-8. Nat Commun. 2025. PMID: 39833157 Free PMC article.
-
Dynamic allostery in the peptide/MHC complex enables TCR neoantigen selectivity.Res Sq [Preprint]. 2024 May 29:rs.3.rs-4457195. doi: 10.21203/rs.3.rs-4457195/v1. Res Sq. 2024. Update in: Nat Commun. 2025 Jan 20;16(1):849. doi: 10.1038/s41467-025-56004-8. PMID: 38854019 Free PMC article. Updated. Preprint.
References
-
- Gross L. Intradermal immunization of C3H mice against a sarcoma that originated in an animal of the same line. Cancer Res. 1943;3(5):326–333.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
