

Minimalist Tetrazine N-Acetyl Muramic Acid Probes for Rapid and Efficient Labeling of Commensal and Pathogenic Peptidoglycans in Living Bacterial Culture and During Macrophage Invasion
- PMID: 38427023
- PMCID: PMC10941766
- DOI: 10.1021/jacs.3c13644
Minimalist Tetrazine N-Acetyl Muramic Acid Probes for Rapid and Efficient Labeling of Commensal and Pathogenic Peptidoglycans in Living Bacterial Culture and During Macrophage Invasion
Abstract
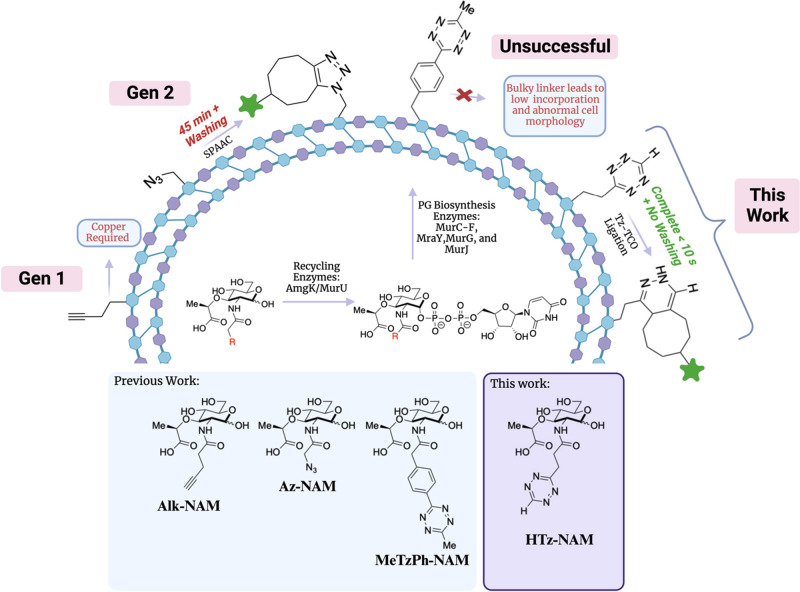
N-Acetyl muramic acid (NAM) probes containing alkyne or azide groups are commonly used to investigate aspects of cell wall synthesis because of their small size and ability to incorporate into bacterial peptidoglycan (PG). However, copper-catalyzed alkyne-azide cycloaddition (CuAAC) reactions are not compatible with live cells, and strain-promoted alkyne-azide cycloaddition (SPAAC) reaction rates are modest and, therefore, not as desirable for tracking the temporal alterations of bacterial cell growth, remodeling, and division. Alternatively, the tetrazine-trans-cyclooctene ligation (Tz-TCO), which is the fastest known bioorthogonal reaction and not cytotoxic, allows for rapid live-cell labeling of PG at biologically relevant time scales and concentrations. Previous work to increase reaction kinetics on the PG surface by using tetrazine probes was limited because of low incorporation of the probe. Described here are new approaches to construct a minimalist tetrazine (Tz)-NAM probe utilizing recent advancements in asymmetric tetrazine synthesis. This minimalist Tz-NAM probe was successfully incorporated into pathogenic and commensal bacterial PG where fixed and rapid live-cell, no-wash labeling was successful in both free bacterial cultures and in coculture with human macrophages. Overall, this probe allows for expeditious labeling of bacterial PG, thereby making it an exceptional tool for monitoring PG biosynthesis for the development of new antibiotic screens. The versatility and selectivity of this probe will allow for real-time interrogation of the interactions of bacterial pathogens in a human host and will serve a broader utility for studying glycans in multiple complex biological systems.
Conflict of interest statement
The authors declare no competing financial interest.
Figures





Similar articles
-
Metabolic Incorporation of N-Acetyl Muramic Acid Probes into Bacterial Peptidoglycan.Curr Protoc Chem Biol. 2019 Dec;11(4):e74. doi: 10.1002/cpch.74. Curr Protoc Chem Biol. 2019. PMID: 31763799 Free PMC article.
-
From mechanism to mouse: a tale of two bioorthogonal reactions.Acc Chem Res. 2011 Sep 20;44(9):666-76. doi: 10.1021/ar200148z. Epub 2011 Aug 15. Acc Chem Res. 2011. PMID: 21838330 Free PMC article.
-
IEDDA: An Attractive Bioorthogonal Reaction for Biomedical Applications.Molecules. 2021 Jul 30;26(15):4640. doi: 10.3390/molecules26154640. Molecules. 2021. PMID: 34361793 Free PMC article. Review.
-
Advances in Tetrazine Bioorthogonal Chemistry Driven by the Synthesis of Novel Tetrazines and Dienophiles.Acc Chem Res. 2018 May 15;51(5):1249-1259. doi: 10.1021/acs.accounts.8b00062. Epub 2018 Apr 11. Acc Chem Res. 2018. PMID: 29638113 Free PMC article.
-
Enzyme- or light-triggered cyclopropenes for bioorthogonal ligation.Methods Enzymol. 2020;641:1-34. doi: 10.1016/bs.mie.2020.04.034. Epub 2020 Jun 12. Methods Enzymol. 2020. PMID: 32713519 Review.
Cited by
-
Bio-orthogonal Labeling of Chitin in Native Pathogenic Candida Species via the Chitin Scavenge Pathway.J Am Chem Soc. 2025 Feb 19;147(7):5632-5641. doi: 10.1021/jacs.4c11554. Epub 2025 Feb 9. J Am Chem Soc. 2025. PMID: 39925016
-
Chemical tools to study and modulate glycan-mediated host-bacteria interactions.Curr Opin Chem Biol. 2025 Aug;87:102603. doi: 10.1016/j.cbpa.2025.102603. Epub 2025 Jun 4. Curr Opin Chem Biol. 2025. PMID: 40472591 Review.
-
Manipulating DNA and RNA structures via click-to-release caged nucleic acids for biological and biomedical applications.Nucleic Acids Res. 2025 Jun 20;53(12):gkaf571. doi: 10.1093/nar/gkaf571. Nucleic Acids Res. 2025. PMID: 40598897 Free PMC article.
-
The Legionella pneumophila peptidoglycan recycling kinase, AmgK, is essential for survival and replication inside host alveolar macrophages.bioRxiv [Preprint]. 2025 Mar 22:2025.03.21.644609. doi: 10.1101/2025.03.21.644609. bioRxiv. 2025. PMID: 40166355 Free PMC article. Preprint.
-
Discovering microbiota functions via chemical probe incorporation for targeted sequencing.Curr Opin Chem Biol. 2025 Feb;84:102551. doi: 10.1016/j.cbpa.2024.102551. Epub 2024 Nov 30. Curr Opin Chem Biol. 2025. PMID: 39615426 Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
