

The CARD8 inflammasome dictates HIV/SIV pathogenesis and disease progression
- PMID: 38428396
- PMCID: PMC10919936
- DOI: 10.1016/j.cell.2024.01.048
The CARD8 inflammasome dictates HIV/SIV pathogenesis and disease progression
Abstract
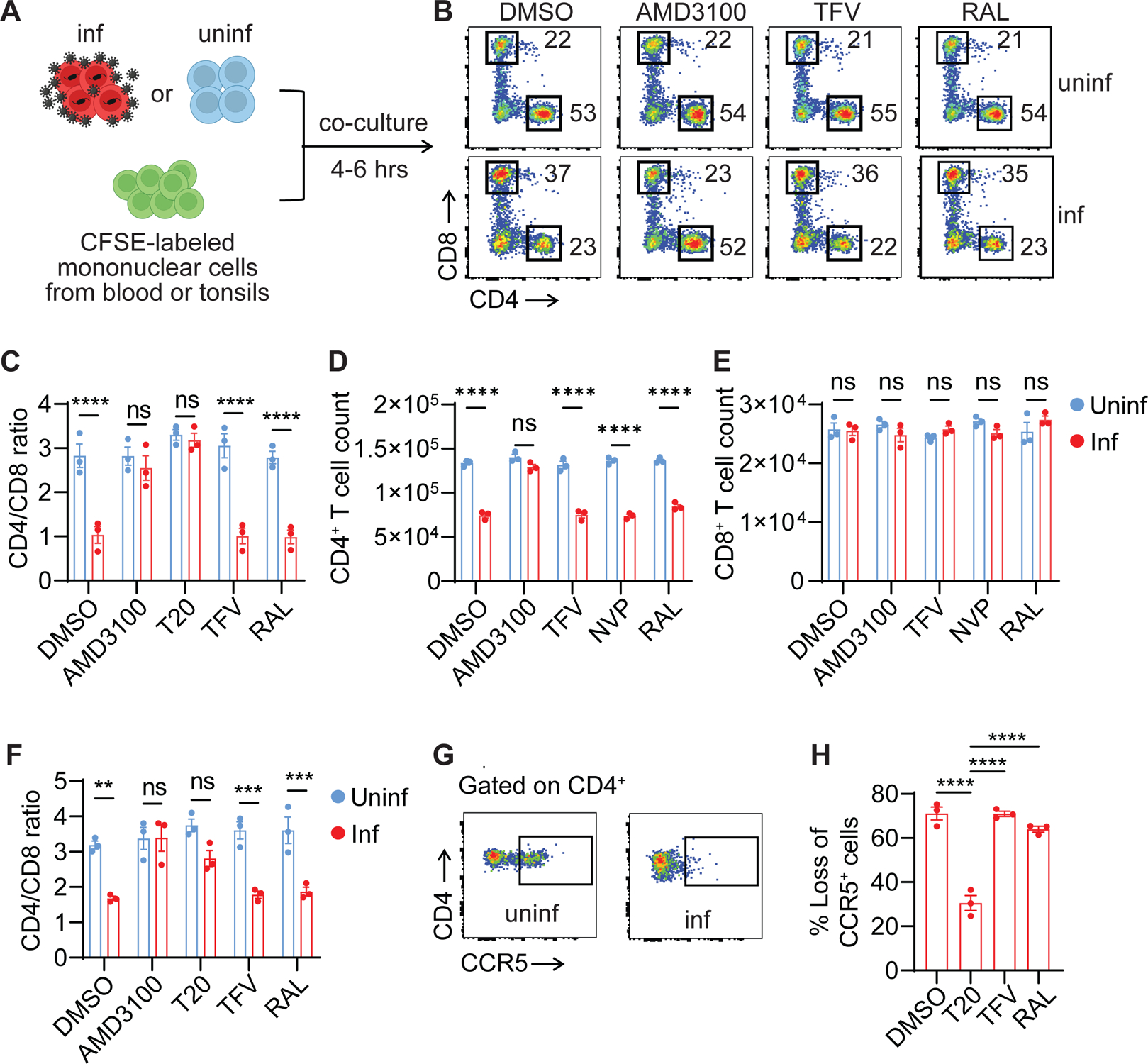
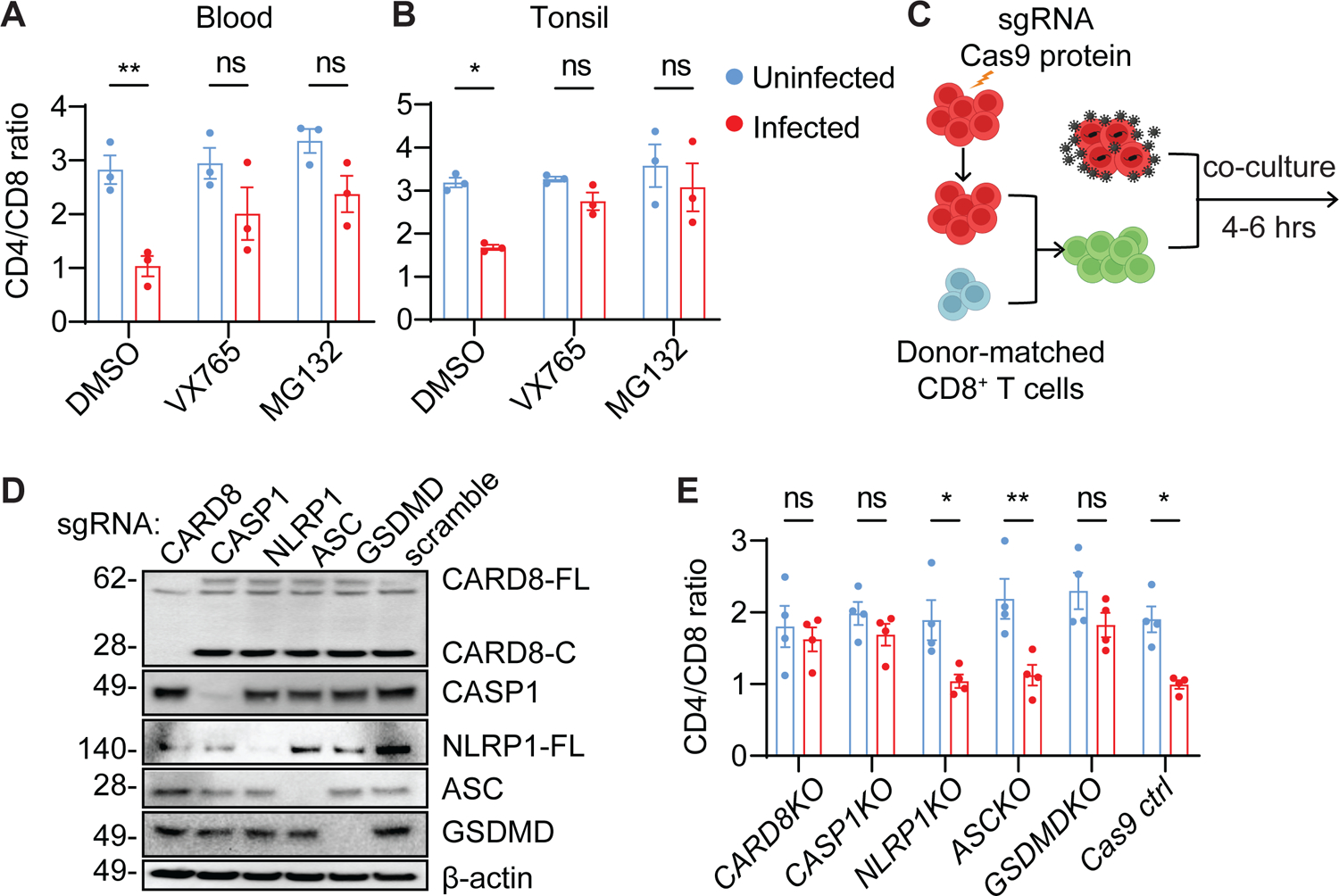
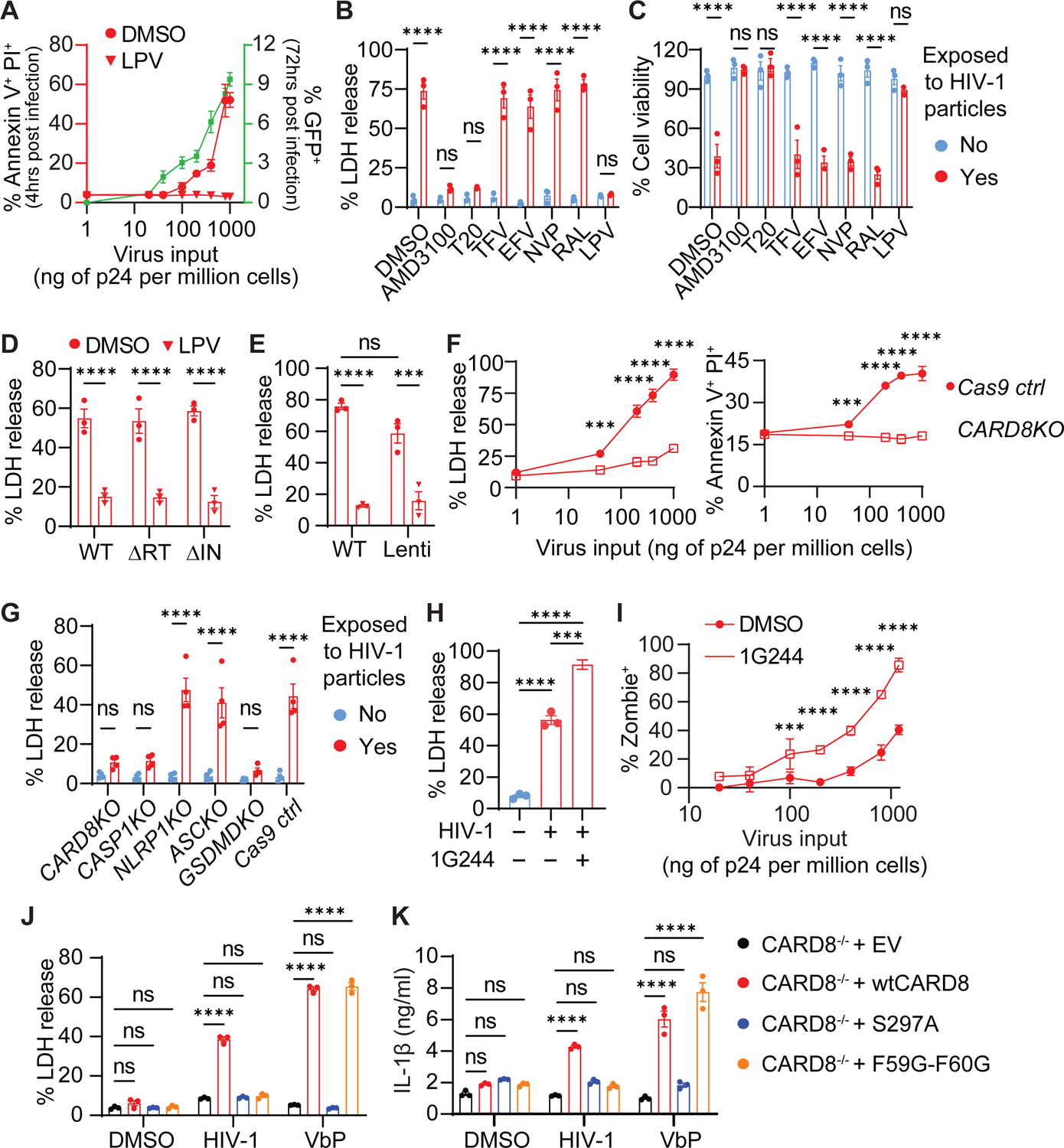
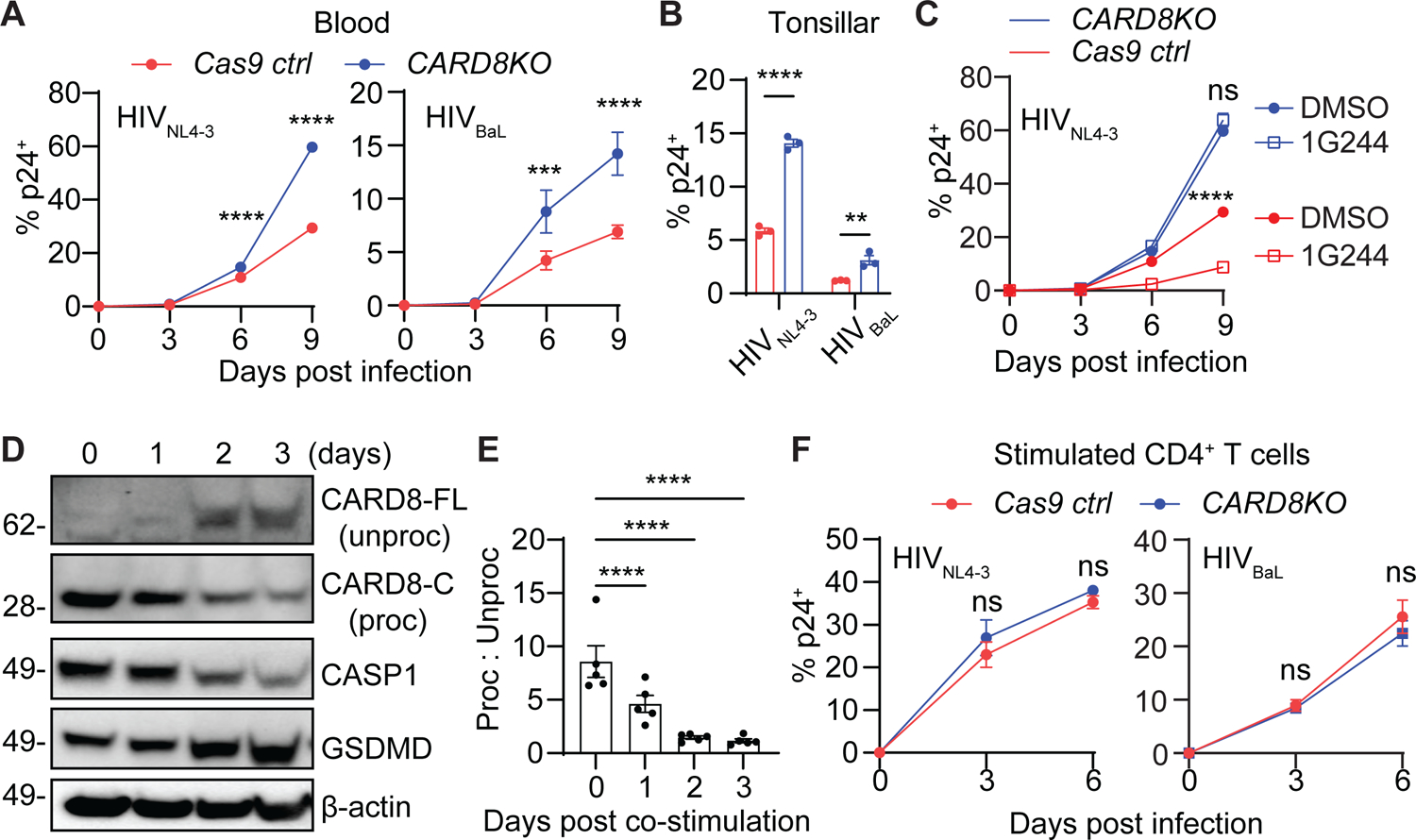
While CD4+ T cell depletion is key to disease progression in people living with HIV and SIV-infected macaques, the mechanisms underlying this depletion remain incompletely understood, with most cell death involving uninfected cells. In contrast, SIV infection of "natural" hosts such as sooty mangabeys does not cause CD4+ depletion and AIDS despite high-level viremia. Here, we report that the CARD8 inflammasome is activated immediately after HIV entry by the viral protease encapsulated in incoming virions. Sensing of HIV protease activity by CARD8 leads to rapid pyroptosis of quiescent cells without productive infection, while T cell activation abolishes CARD8 function and increases permissiveness to infection. In humanized mice reconstituted with CARD8-deficient cells, CD4+ depletion is delayed despite high viremia. Finally, we discovered loss-of-function mutations in CARD8 from "natural hosts," which may explain the peculiarly non-pathogenic nature of these infections. Our study suggests that CARD8 drives CD4+ T cell depletion during pathogenic HIV/SIV infections.
Keywords: CARD8; CD4(+) T cells; HIV; SIV; caspase 1; inflammasome; non-human primates; pyroptosis; viral entry; viral protease.
Copyright © 2024 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
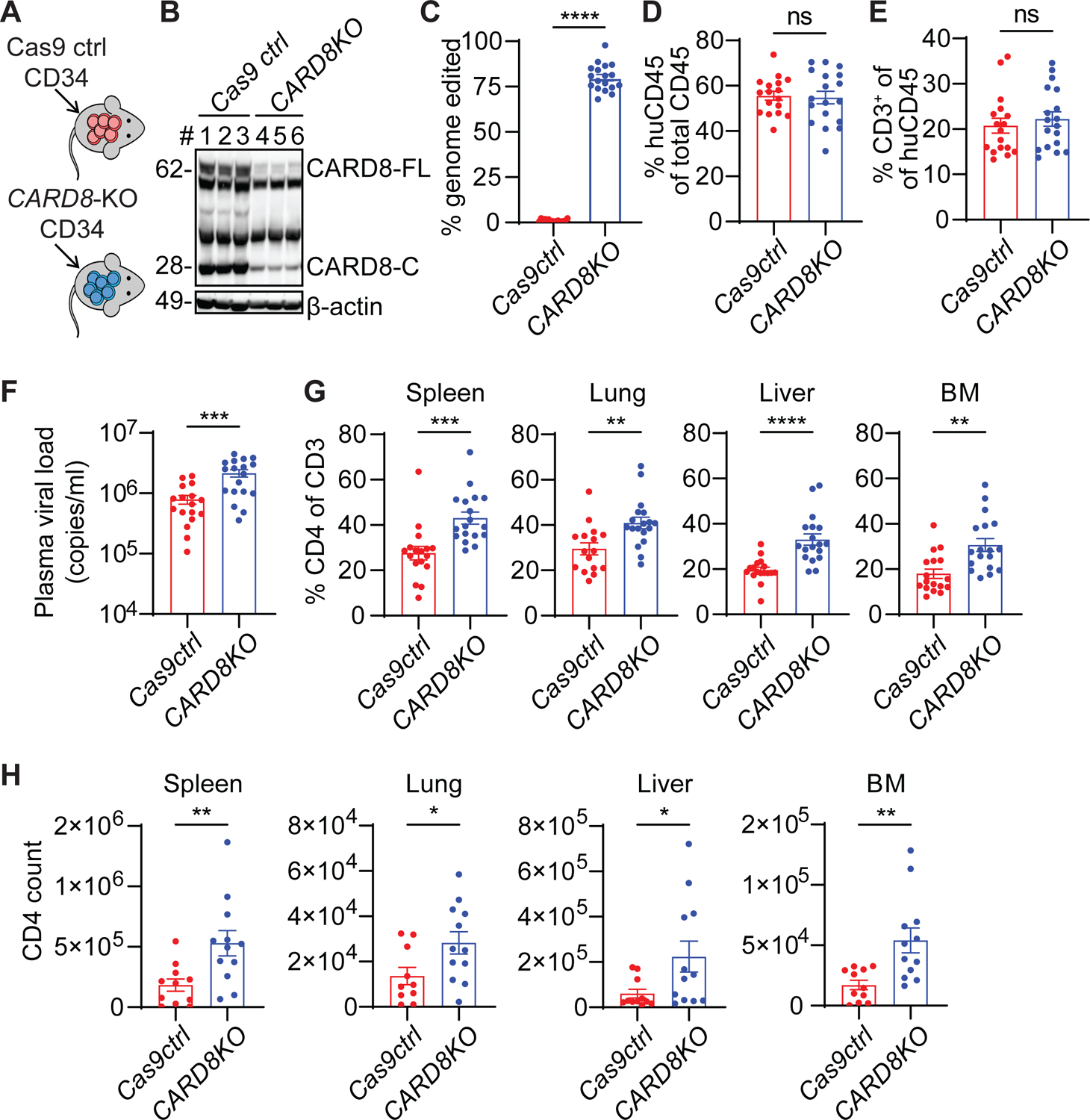
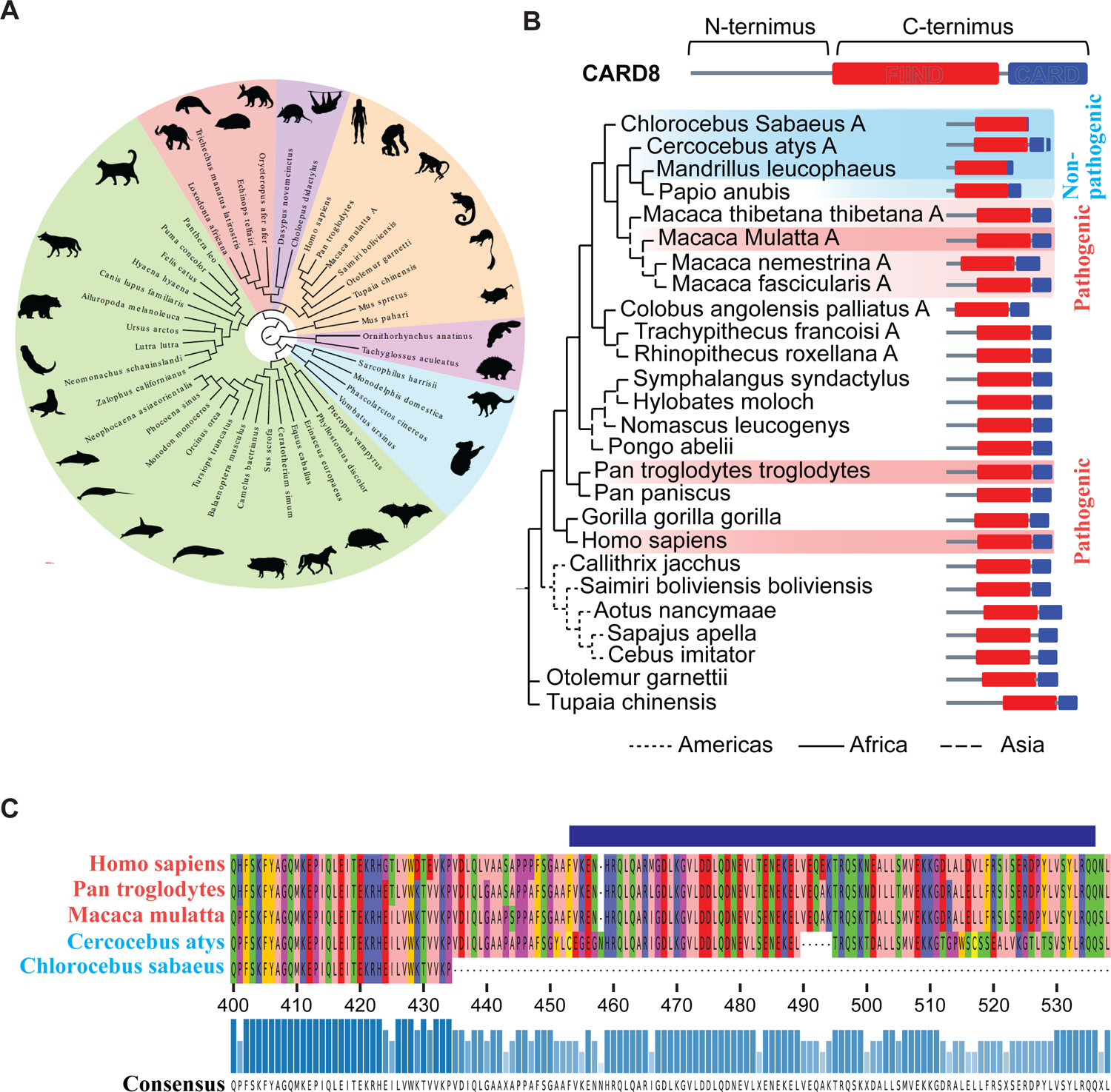
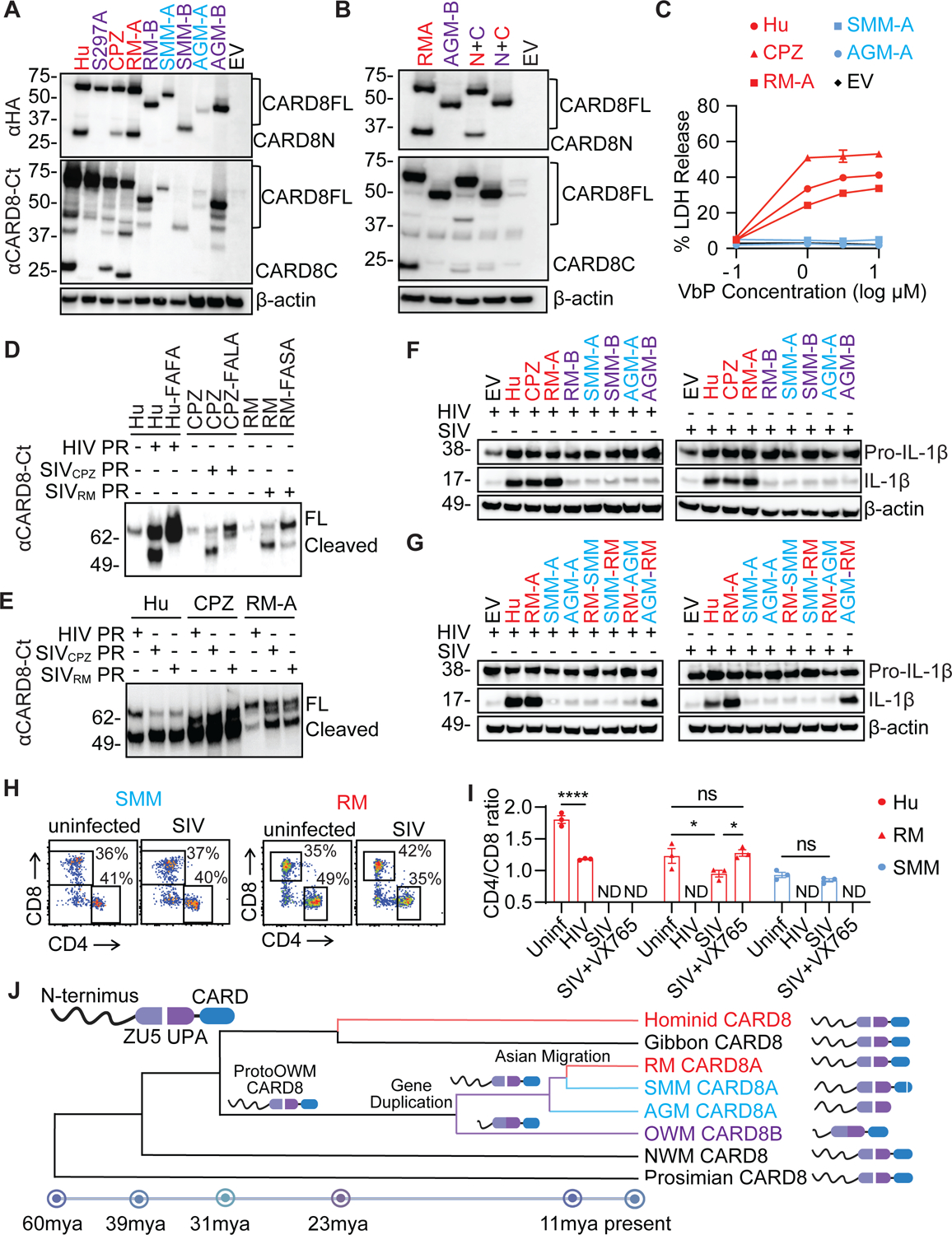
References
-
- Psallidopoulos MC, Schnittman SM, Thompson LM 3rd, Baseler M, Fauci AS, Lane HC, and Salzman NP (1989). Integrated proviral human immunodeficiency virus type 1 is present in CD4+ peripheral blood lymphocytes in healthy seropositive individuals. J Virol 63, 4626–4631. 10.1128/JVI.63.11.4626-4631.1989. - DOI - PMC - PubMed
-
- Schnittman SM, Greenhouse JJ, Psallidopoulos MC, Baseler M, Salzman NP, Fauci AS, and Lane HC (1990). Increasing viral burden in CD4+ T cells from patients with human immunodeficiency virus (HIV) infection reflects rapidly progressive immunosuppression and clinical disease. Ann Intern Med 113, 438–443. 10.7326/0003-4819-113-6-438. - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
