

Structurally divergent and recurrently mutated regions of primate genomes
- PMID: 38428424
- PMCID: PMC10947866
- DOI: 10.1016/j.cell.2024.01.052
Structurally divergent and recurrently mutated regions of primate genomes
Abstract
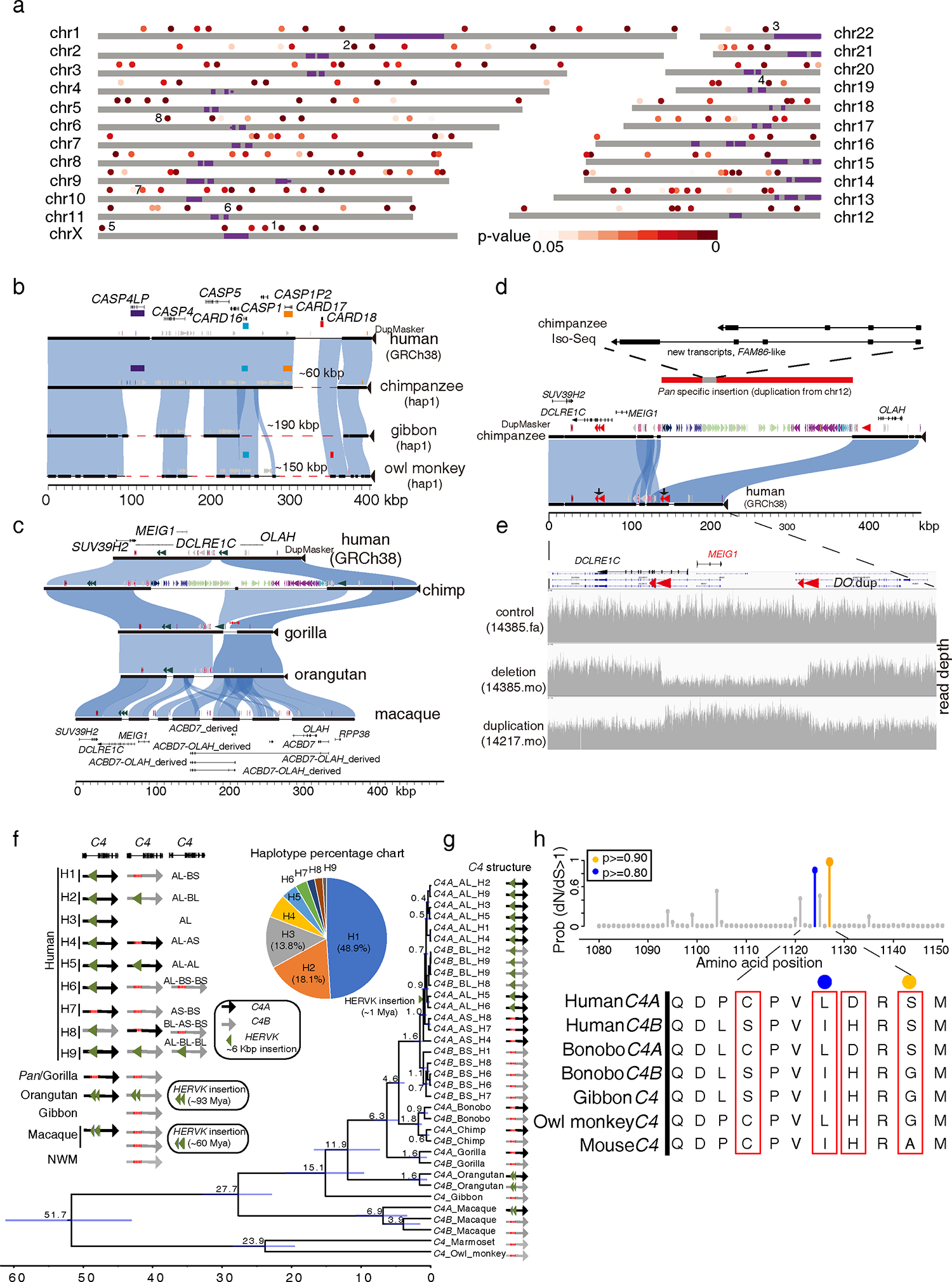
We sequenced and assembled using multiple long-read sequencing technologies the genomes of chimpanzee, bonobo, gorilla, orangutan, gibbon, macaque, owl monkey, and marmoset. We identified 1,338,997 lineage-specific fixed structural variants (SVs) disrupting 1,561 protein-coding genes and 136,932 regulatory elements, including the most complete set of human-specific fixed differences. We estimate that 819.47 Mbp or ∼27% of the genome has been affected by SVs across primate evolution. We identify 1,607 structurally divergent regions wherein recurrent structural variation contributes to creating SV hotspots where genes are recurrently lost (e.g., CARD, C4, and OLAH gene families) and additional lineage-specific genes are generated (e.g., CKAP2, VPS36, ACBD7, and NEK5 paralogs), becoming targets of rapid chromosomal diversification and positive selection (e.g., RGPD gene family). High-fidelity long-read sequencing has made these dynamic regions of the genome accessible for sequence-level analyses within and between primate species.
Keywords: NPHP1 and Joubert syndrome; RGPD gene family; adaptive evolution; comparative genomics; duplicated genes; evolutionary medicine; human diseases; long-read sequencing; primate evolution.
Copyright © 2024 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests E.E.E. is a scientific advisory board (SAB) member of Variant Bio, Inc.
Figures




Update of
-
Structurally divergent and recurrently mutated regions of primate genomes.bioRxiv [Preprint]. 2023 Mar 7:2023.03.07.531415. doi: 10.1101/2023.03.07.531415. bioRxiv. 2023. Update in: Cell. 2024 Mar 14;187(6):1547-1562.e13. doi: 10.1016/j.cell.2024.01.052. PMID: 36945442 Free PMC article. Updated. Preprint.
References
-
- Lander ES, Linton LM, Birren B, Nusbaum C, Zody MC, Baldwin J, Devon K, Dewar K, Doyle M, FitzHugh W, et al. (2001). Initial sequencing and analysis of the human genome. Nature 409, 860–921. - PubMed
-
- Venter JC, Adams MD, Myers EW, Li PW, Mural RJ, Sutton GG, Smith HO, Yandell M, Evans CA, Holt RA et al. (2001). The sequence of the human genome. Science 291, 1304–1351. - PubMed
-
- Watson JD (1990). The human genome project: past, present, and future. Science 248, 44–49. - PubMed
MeSH terms
Grants and funding
- DP1 AI175471/AI/NIAID NIH HHS/United States
- U41 HG010972/HG/NHGRI NIH HHS/United States
- R01 AI137011/AI/NIAID NIH HHS/United States
- K99 GM147352/GM/NIGMS NIH HHS/United States
- U24 HG011853/HG/NHGRI NIH HHS/United States
- R01 HG010333/HG/NHGRI NIH HHS/United States
- R01 HG010485/HG/NHGRI NIH HHS/United States
- R01 OD034046/OD/NIH HHS/United States
- R01 HG002385/HG/NHGRI NIH HHS/United States
- P51 OD011092/OD/NIH HHS/United States
- U01 HG010961/HG/NHGRI NIH HHS/United States
- DP1 DA046108/DA/NIDA NIH HHS/United States
- R00 GM147352/GM/NIGMS NIH HHS/United States
- R01 HG010169/HG/NHGRI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Miscellaneous
