

AutoRNC: An automated modeling program for building atomic models of ribosome-nascent chain complexes
- PMID: 38428431
- PMCID: PMC11073581
- DOI: 10.1016/j.str.2024.02.002
AutoRNC: An automated modeling program for building atomic models of ribosome-nascent chain complexes
Abstract
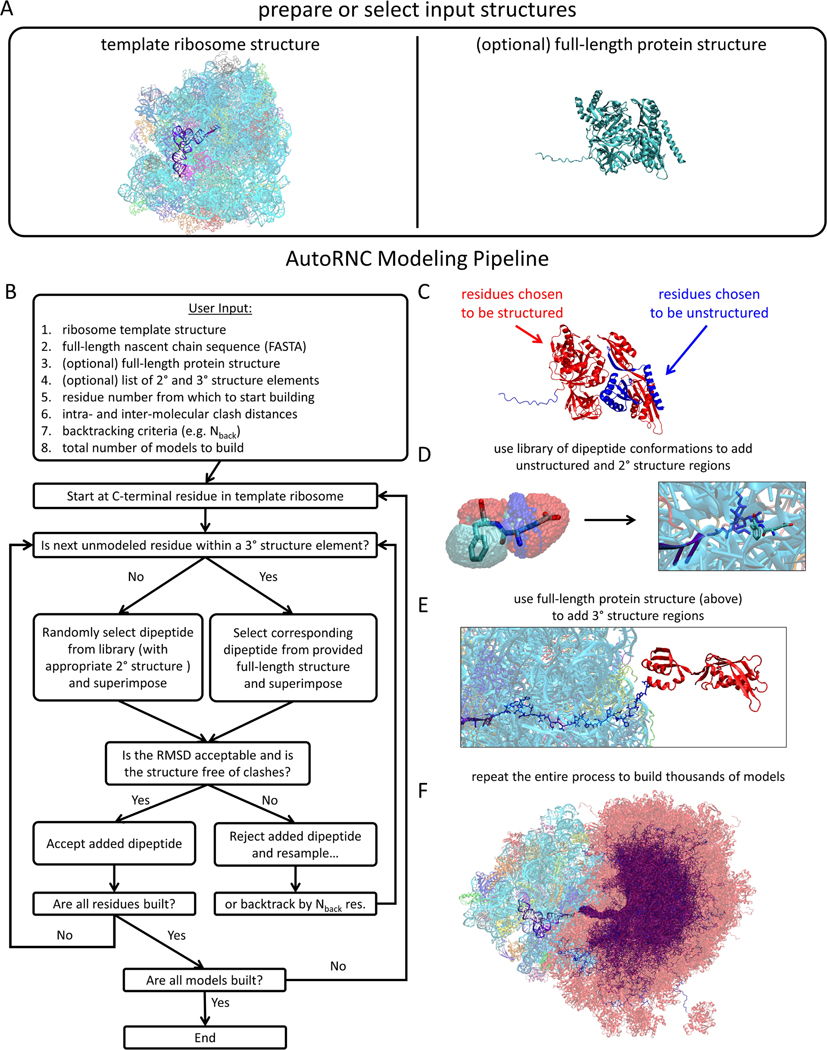
The interpretation of experimental studies of co-translational protein folding often benefits from the use of computational methods that seek to model or simulate the nascent chain and its interactions with the ribosome. Building realistic 3D models of ribosome-nascent chain (RNC) constructs often requires expert knowledge, so to circumvent this issue, we describe here AutoRNC, an automated modeling program capable of constructing large numbers of plausible atomic models of RNCs within minutes. AutoRNC takes input from the user specifying any regions of the nascent chain that contain secondary or tertiary structure and attempts to build conformations compatible with those specifications-and with the constraints imposed by the ribosome-by sampling and progressively piecing together dipeptide conformations extracted from the Research Collaboratory for Structural Bioinformatics Protein Data Bank (RCSB). Despite using only modest computational resources, we show here that AutoRNC can build plausible conformations for a wide range of RNC constructs for which experimental data have already been reported.
Copyright © 2024 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures



Update of
-
AutoRNC: an automated modeling program for building atomic models of ribosome-nascent chain complexes.bioRxiv [Preprint]. 2023 Jun 14:2023.06.14.544999. doi: 10.1101/2023.06.14.544999. bioRxiv. 2023. Update in: Structure. 2024 May 2;32(5):621-629.e5. doi: 10.1016/j.str.2024.02.002. PMID: 37398297 Free PMC article. Updated. Preprint.
Similar articles
-
AutoRNC: an automated modeling program for building atomic models of ribosome-nascent chain complexes.bioRxiv [Preprint]. 2023 Jun 14:2023.06.14.544999. doi: 10.1101/2023.06.14.544999. bioRxiv. 2023. Update in: Structure. 2024 May 2;32(5):621-629.e5. doi: 10.1016/j.str.2024.02.002. PMID: 37398297 Free PMC article. Updated. Preprint.
-
Cost-effectiveness of using prognostic information to select women with breast cancer for adjuvant systemic therapy.Health Technol Assess. 2006 Sep;10(34):iii-iv, ix-xi, 1-204. doi: 10.3310/hta10340. Health Technol Assess. 2006. PMID: 16959170
-
Signs and symptoms to determine if a patient presenting in primary care or hospital outpatient settings has COVID-19.Cochrane Database Syst Rev. 2022 May 20;5(5):CD013665. doi: 10.1002/14651858.CD013665.pub3. Cochrane Database Syst Rev. 2022. PMID: 35593186 Free PMC article.
-
Technological aids for the rehabilitation of memory and executive functioning in children and adolescents with acquired brain injury.Cochrane Database Syst Rev. 2016 Jul 1;7(7):CD011020. doi: 10.1002/14651858.CD011020.pub2. Cochrane Database Syst Rev. 2016. PMID: 27364851 Free PMC article.
-
Nivolumab for adults with Hodgkin's lymphoma (a rapid review using the software RobotReviewer).Cochrane Database Syst Rev. 2018 Jul 12;7(7):CD012556. doi: 10.1002/14651858.CD012556.pub2. Cochrane Database Syst Rev. 2018. PMID: 30001476 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
