

Acetylcholine Engages Distinct Amygdala Microcircuits to Gate Internal Theta Rhythm
- PMID: 38438258
- PMCID: PMC11055655
- DOI: 10.1523/JNEUROSCI.1568-23.2024
Acetylcholine Engages Distinct Amygdala Microcircuits to Gate Internal Theta Rhythm
Abstract
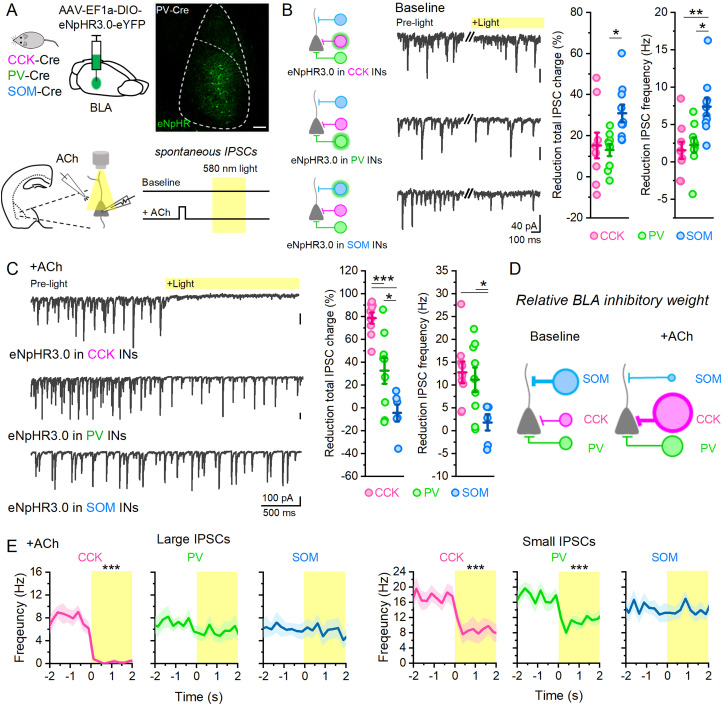
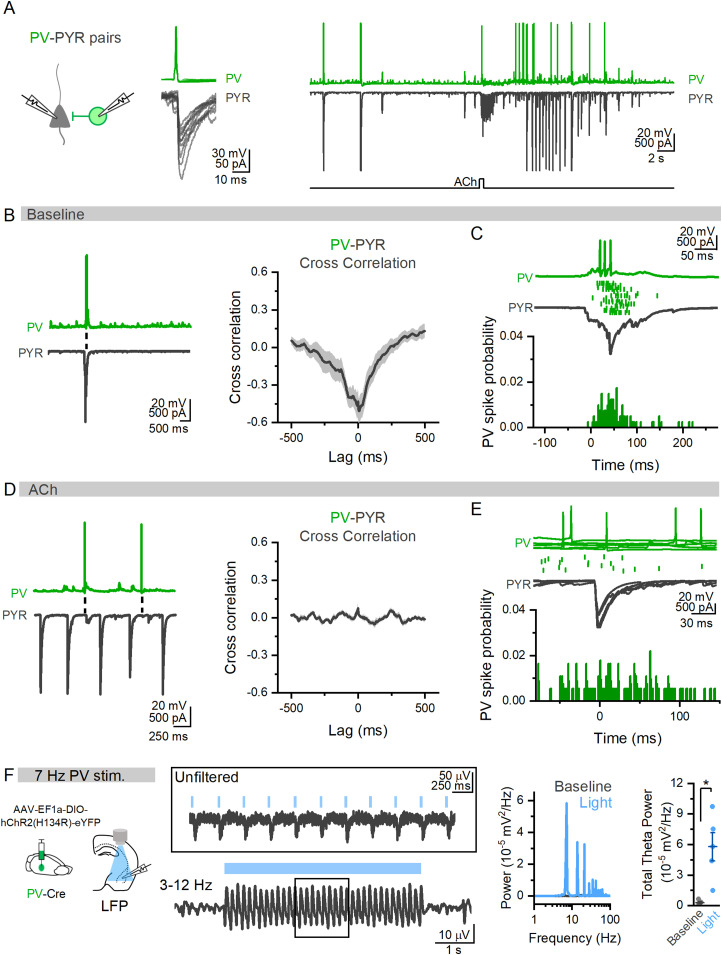
Acetylcholine (ACh) is released from basal forebrain cholinergic neurons in response to salient stimuli and engages brain states supporting attention and memory. These high ACh states are associated with theta oscillations, which synchronize neuronal ensembles. Theta oscillations in the basolateral amygdala (BLA) in both humans and rodents have been shown to underlie emotional memory, yet their mechanism remains unclear. Here, using brain slice electrophysiology in male and female mice, we show large ACh stimuli evoke prolonged theta oscillations in BLA local field potentials that depend upon M3 muscarinic receptor activation of cholecystokinin (CCK) interneurons (INs) without the need for external glutamate signaling. Somatostatin (SOM) INs inhibit CCK INs and are themselves inhibited by ACh, providing a functional SOM→CCK IN circuit connection gating BLA theta. Parvalbumin (PV) INs, which can drive BLA oscillations in baseline states, are not involved in the generation of ACh-induced theta, highlighting that ACh induces a cellular switch in the control of BLA oscillatory activity and establishes an internally BLA-driven theta oscillation through CCK INs. Theta activity is more readily evoked in BLA over the cortex or hippocampus, suggesting preferential activation of the BLA during high ACh states. These data reveal a SOM→CCK IN circuit in the BLA that gates internal theta oscillations and suggest a mechanism by which salient stimuli acting through ACh switch the BLA into a network state enabling emotional memory.
Keywords: acetylcholine; amygdala; emotion; interneuron; oscillation; theta.
Copyright © 2024 the authors.
Conflict of interest statement
The authors declare no competing financial interests.
Figures









References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases