

Regulation of developmental gatekeeping and cell fate transition by the calpain protease DEK1 in Physcomitrium patens
- PMID: 38438476
- PMCID: PMC10912778
- DOI: 10.1038/s42003-024-05933-z
Regulation of developmental gatekeeping and cell fate transition by the calpain protease DEK1 in Physcomitrium patens
Abstract
Calpains are cysteine proteases that control cell fate transitions whose loss of function causes severe, pleiotropic phenotypes in eukaryotes. Although mainly considered as modulatory proteases, human calpain targets are directed to the N-end rule degradation pathway. Several such targets are transcription factors, hinting at a gene-regulatory role. Here, we analyze the gene-regulatory networks of the moss Physcomitrium patens and characterize the regulons that are misregulated in mutants of the calpain DEFECTIVE KERNEL1 (DEK1). Predicted cleavage patterns of the regulatory hierarchies in five DEK1-controlled subnetworks are consistent with a pleiotropic and regulatory role during cell fate transitions targeting multiple functions. Network structure suggests DEK1-gated sequential transitions between cell fates in 2D-to-3D development. Our method combines comprehensive phenotyping, transcriptomics and data science to dissect phenotypic traits, and our model explains the protease function as a switch gatekeeping cell fate transitions potentially also beyond plant development.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
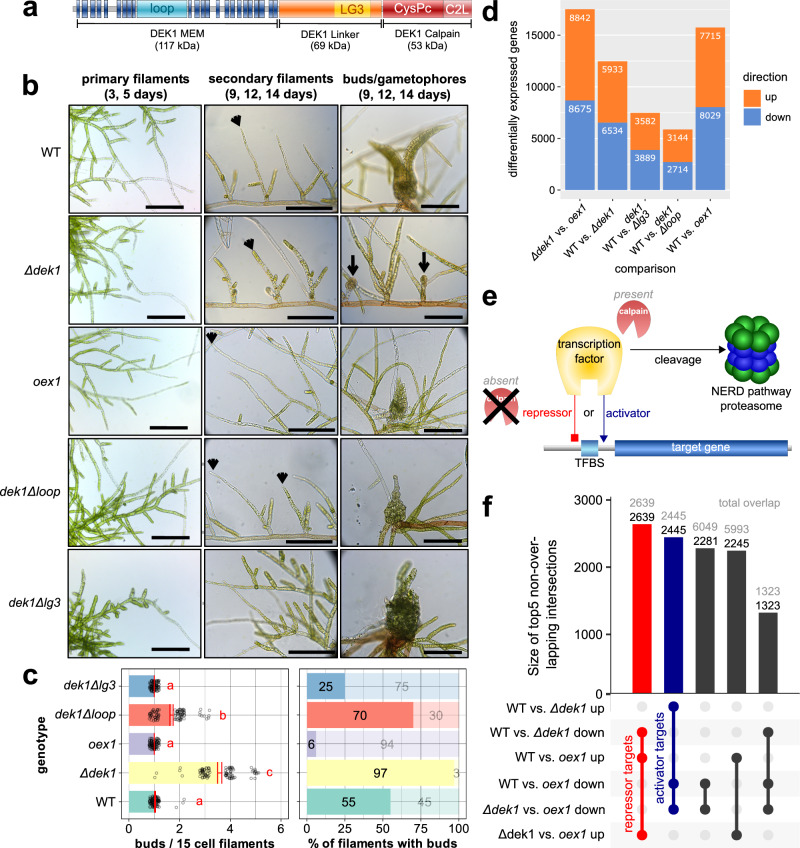
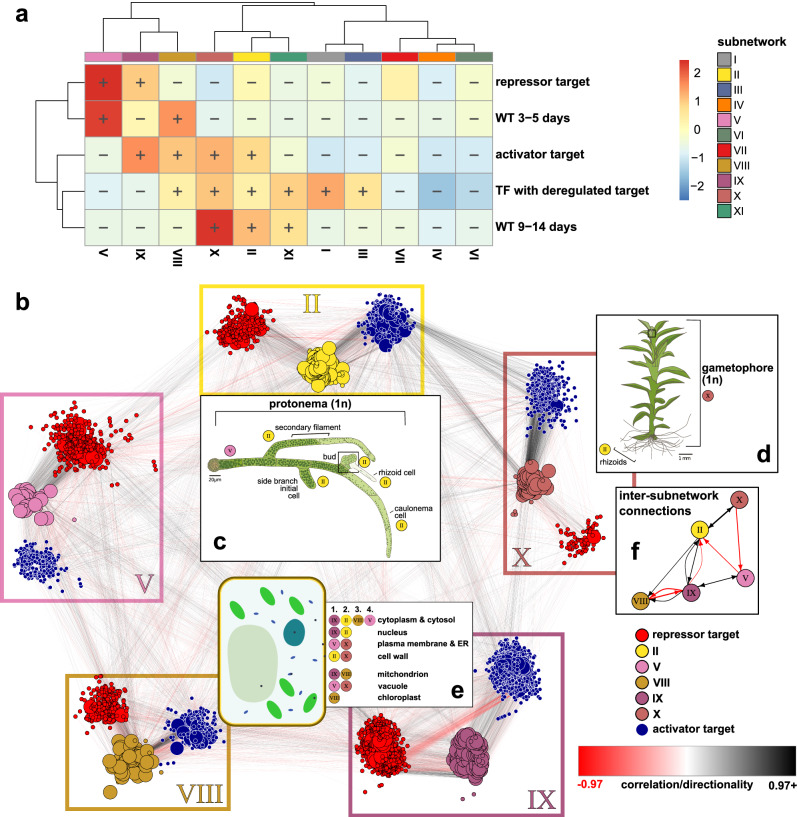
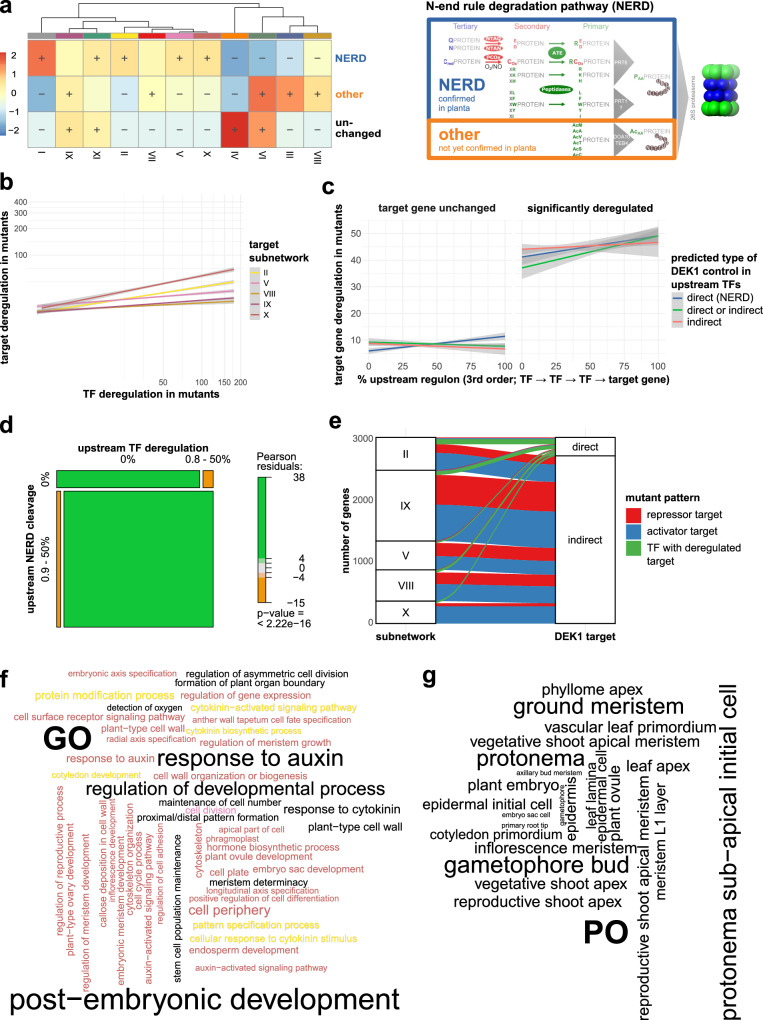
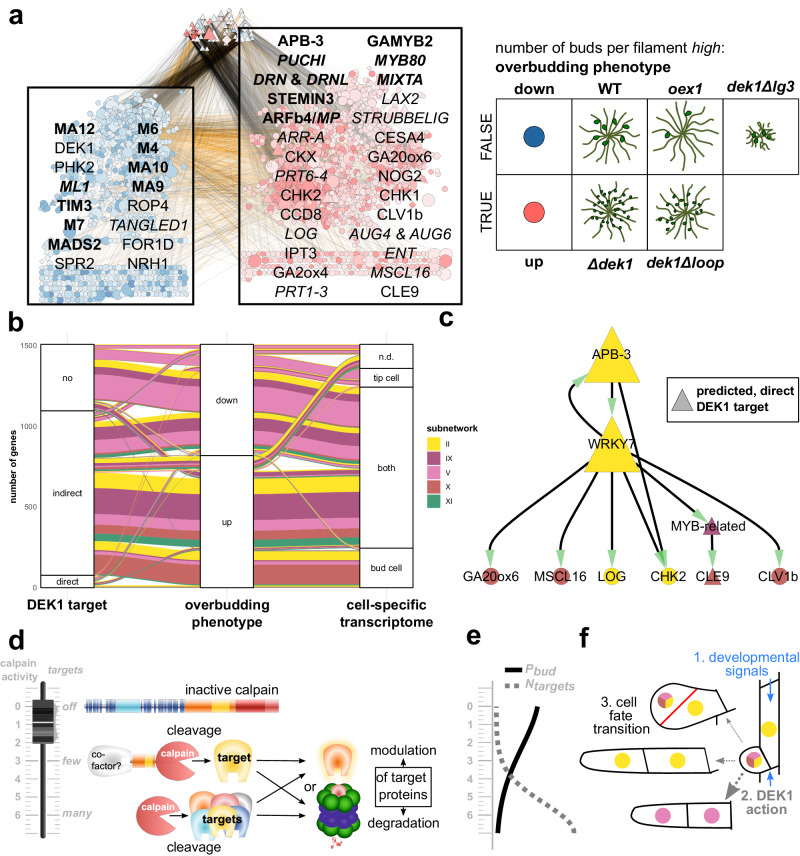
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
