

An array of signal-specific MoYpd1 isoforms determines full virulence in the pathogenic fungus Magnaporthe oryzae
- PMID: 38438487
- PMCID: PMC10912366
- DOI: 10.1038/s42003-024-05941-z
An array of signal-specific MoYpd1 isoforms determines full virulence in the pathogenic fungus Magnaporthe oryzae
Abstract
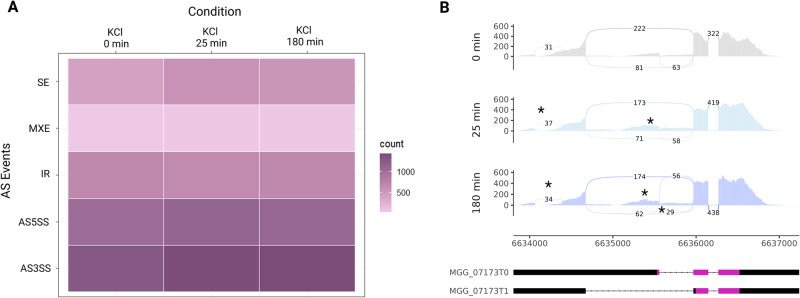
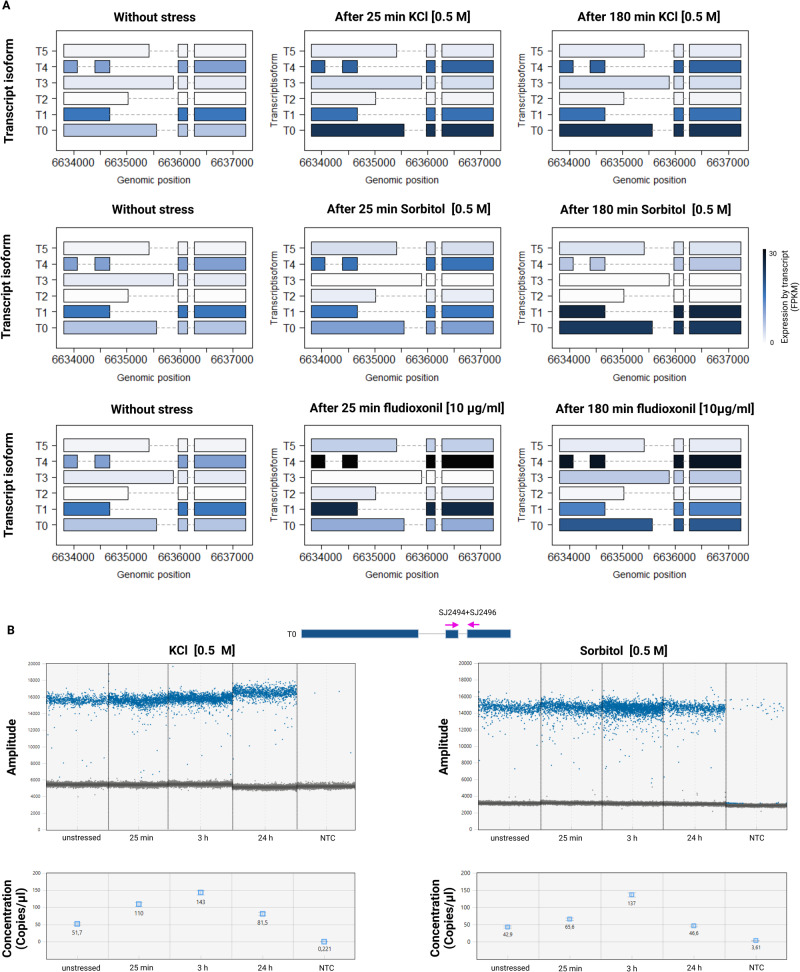
Magnaporthe oryzae is placed first on a list of the world's top ten plant pathogens with the highest scientific and economic importance. The locus MGG_07173 occurs only once in the genome of M. oryzae and encodes the phosphotransfer protein MoYpd1p, which plays an important role in the high osmolarity glycerol (HOG) signaling pathway for osmoregulation. Originating from this locus, at least three MoYPD1 isoforms are produced in a signal-specific manner. The transcript levels of these MoYPD1-isoforms were individually affected by external stress. Salt (KCI) stress raised MoYPD1_T0 abundance, whereas osmotic stress by sorbitol elevates MoYPD1_T1 levels. In line with this, signal-specific nuclear translocation of green fluorescent protein-fused MoYpd1p isoforms in response to stress was observed. Mutant strains that produce only one of the MoYpd1p isoforms are less virulent, suggesting a combination thereof is required to invade the host successfully. In summary, we demonstrate signal-specific production of MoYpd1p isoforms that individually increase signal diversity and orchestrate virulence in M. oryzae.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures




Similar articles
-
High osmolarity glycerol (HOG) signalling in Magnaporthe oryzae: Identification of MoYPD1 and its role in osmoregulation, fungicide action, and pathogenicity.Fungal Biol. 2015 Jul;119(7):580-94. doi: 10.1016/j.funbio.2015.03.003. Epub 2015 Mar 14. Fungal Biol. 2015. PMID: 26058534
-
Evidence of a New MoYpd1p Phosphotransferase Isoform in the Multistep Phosphorelay System of Magnaporthe oryzae.J Fungi (Basel). 2021 May 15;7(5):389. doi: 10.3390/jof7050389. J Fungi (Basel). 2021. PMID: 34063560 Free PMC article.
-
Controllable Bypass Suppression in Magnaporthe oryzae.Methods Mol Biol. 2021;2356:225-231. doi: 10.1007/978-1-0716-1613-0_18. Methods Mol Biol. 2021. PMID: 34236690
-
Regulation of Autophagy Machinery in Magnaporthe oryzae.Int J Mol Sci. 2022 Jul 28;23(15):8366. doi: 10.3390/ijms23158366. Int J Mol Sci. 2022. PMID: 35955497 Free PMC article. Review.
-
Current understanding of HOG-MAPK pathway in Aspergillus fumigatus.Mycopathologia. 2013 Feb;175(1-2):13-23. doi: 10.1007/s11046-012-9600-5. Epub 2012 Nov 18. Mycopathologia. 2013. PMID: 23161019 Review.
References
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
