

In-cell NMR suggests that DNA i-motif levels are strongly depleted in living human cells
- PMID: 38443388
- PMCID: PMC10914786
- DOI: 10.1038/s41467-024-46221-y
In-cell NMR suggests that DNA i-motif levels are strongly depleted in living human cells
Abstract
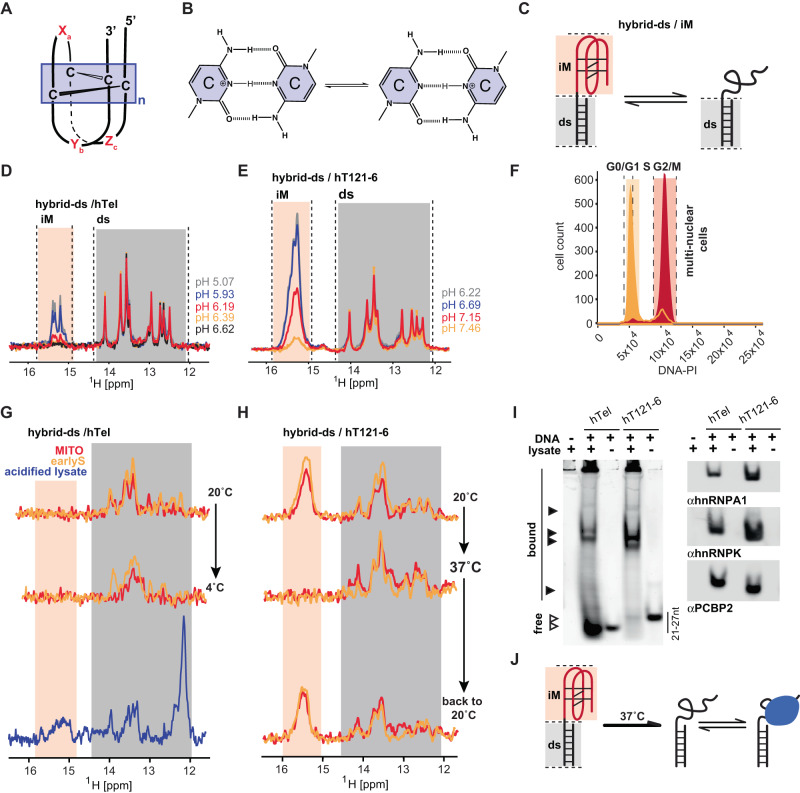
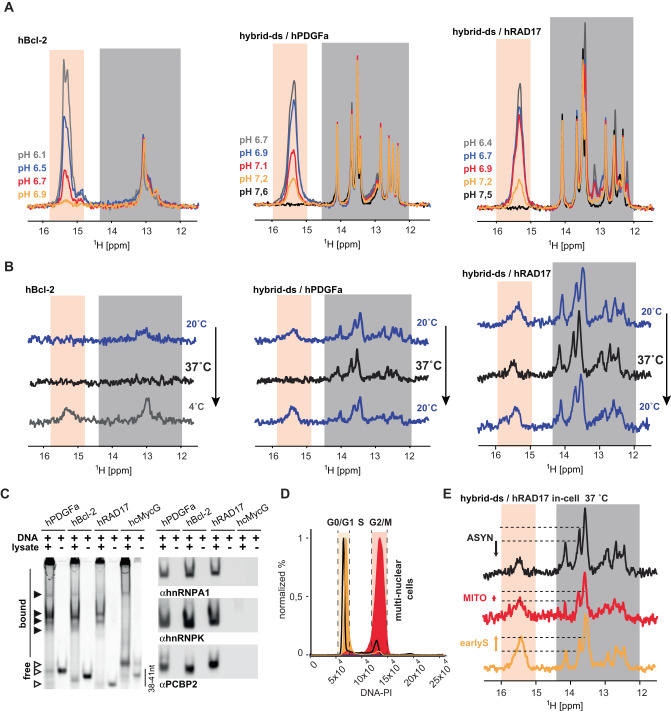
I-Motifs (iM) are non-canonical DNA structures potentially forming in the accessible, single-stranded, cytosine-rich genomic regions with regulatory roles. Chromatin, protein interactions, and intracellular properties seem to govern iM formation at sites with i-motif formation propensity (iMFPS) in human cells, yet their specific contributions remain unclear. Using in-cell NMR with oligonucleotide iMFPS models, we monitor iM-associated structural equilibria in asynchronous and cell cycle-synchronized HeLa cells at 37 °C. Our findings show that iMFPS displaying pHT < 7 under reference in vitro conditions occur predominantly in unfolded states in cells, while those with pHT > 7 appear as a mix of folded and unfolded states depending on the cell cycle phase. Comparing these results with previous data obtained using an iM-specific antibody (iMab) reveals that cell cycle-dependent iM formation has a dual origin, and iM formation concerns only a tiny fraction (possibly 1%) of genomic sites with iM formation propensity. We propose a comprehensive model aligning observations from iMab and in-cell NMR and enabling the identification of iMFPS capable of adopting iM structures under physiological conditions in living human cells. Our results suggest that many iMFPS may have biological roles linked to their unfolded states.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
MeSH terms
Substances
Grants and funding
- CZ.02.2.69/0.0/0.0/20_079/0017045/Ministerstvo Školství, Mládeže a Tělovýchovy (Ministry of Education, Youth and Sports)
- CZ.02.1.01/0.0/0.0/15_003/0000477/Ministerstvo Školství, Mládeže a Tělovýchovy (Ministry of Education, Youth and Sports)
- Discovery Grant/Gouvernement du Canada | Natural Sciences and Engineering Research Council of Canada (Conseil de Recherches en Sciences Naturelles et en Génie du Canada)
- ANR-21-CE44-0005-01/Agence Nationale de la Recherche (French National Research Agency)
LinkOut - more resources
Full Text Sources
Medical
