

Dynamic enhancer landscapes in human craniofacial development
- PMID: 38448444
- PMCID: PMC10917818
- DOI: 10.1038/s41467-024-46396-4
Dynamic enhancer landscapes in human craniofacial development
Abstract
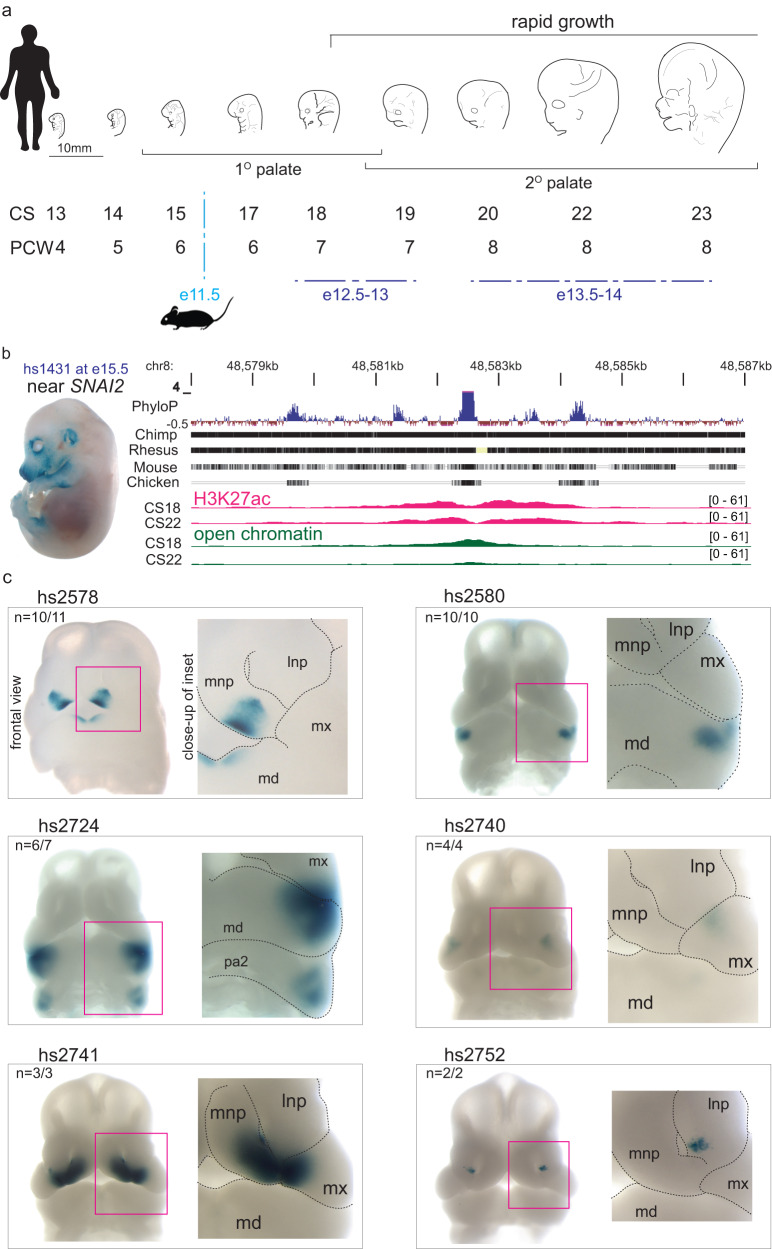
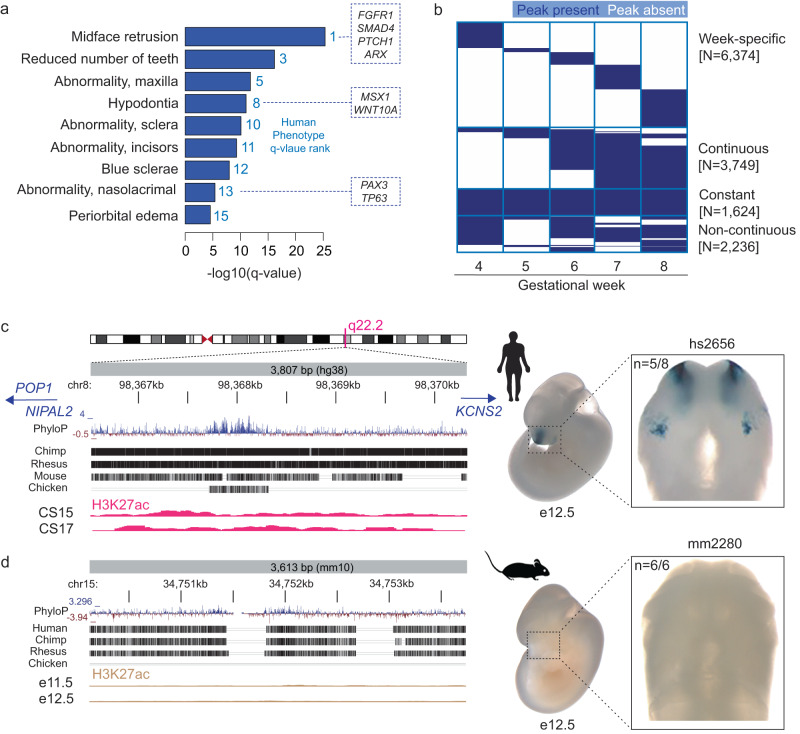
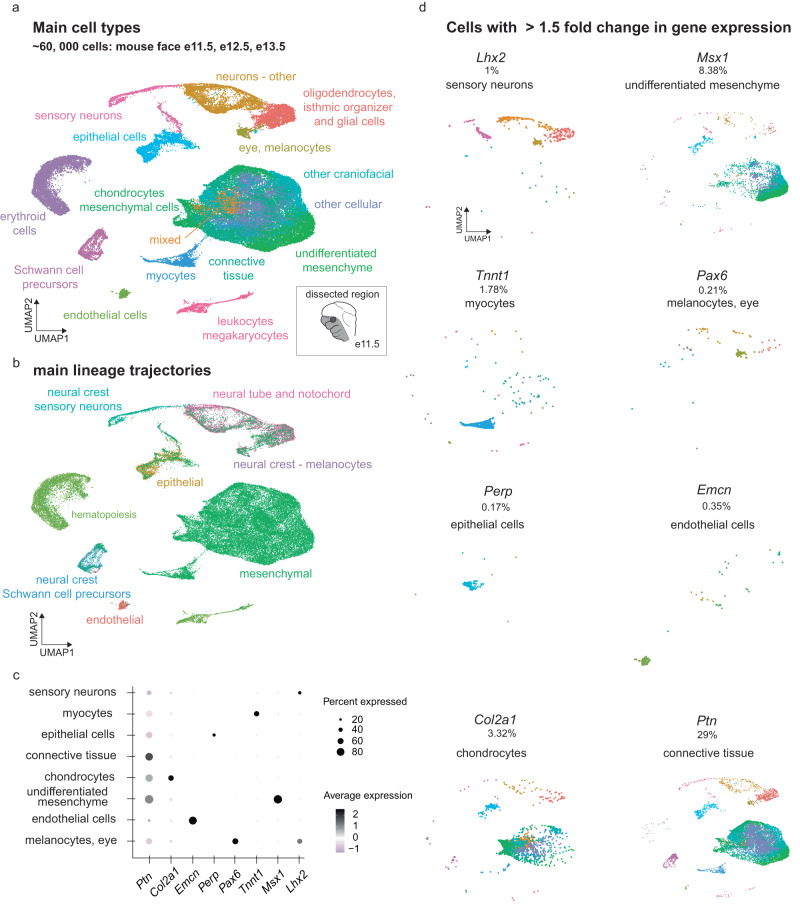
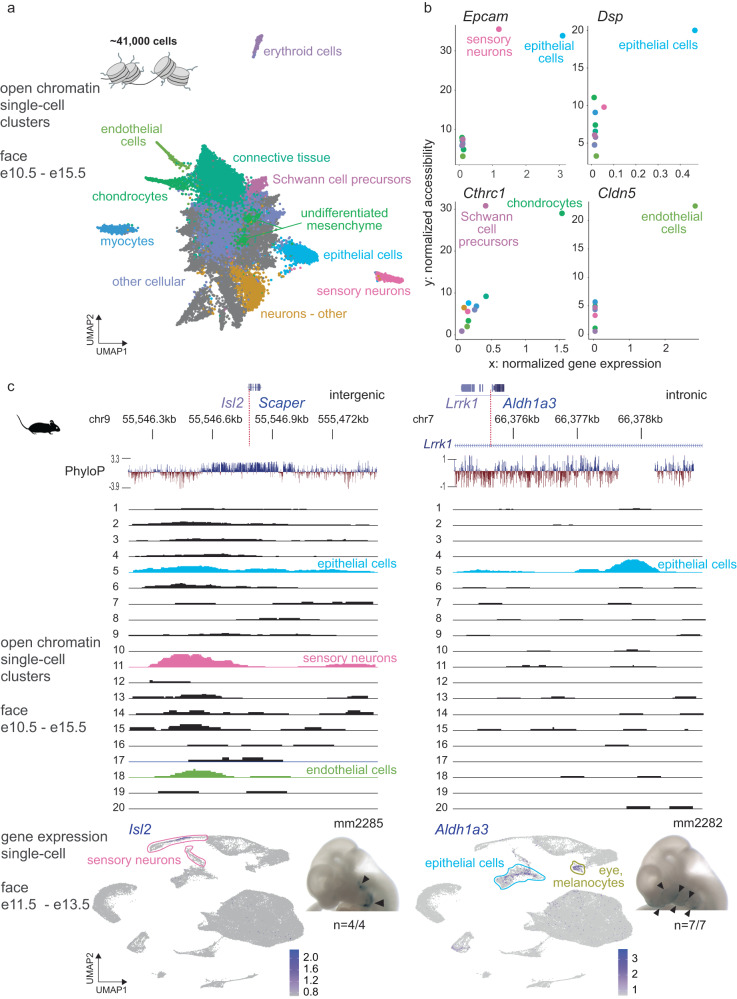
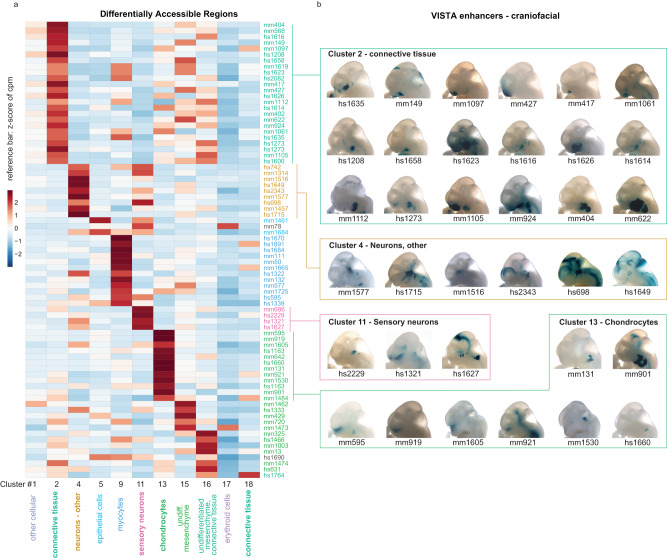
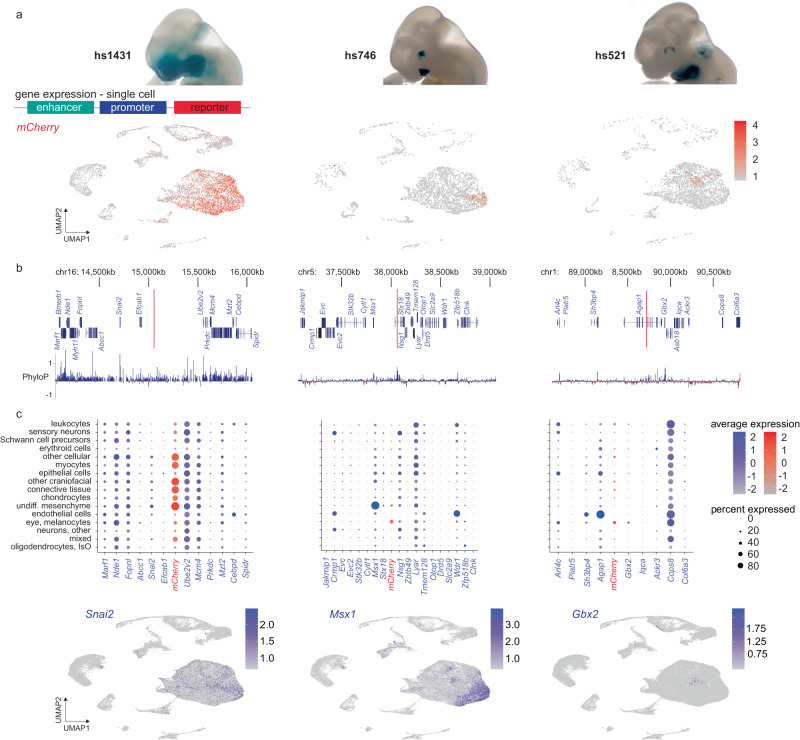
The genetic basis of human facial variation and craniofacial birth defects remains poorly understood. Distant-acting transcriptional enhancers control the fine-tuned spatiotemporal expression of genes during critical stages of craniofacial development. However, a lack of accurate maps of the genomic locations and cell type-resolved activities of craniofacial enhancers prevents their systematic exploration in human genetics studies. Here, we combine histone modification, chromatin accessibility, and gene expression profiling of human craniofacial development with single-cell analyses of the developing mouse face to define the regulatory landscape of facial development at tissue- and single cell-resolution. We provide temporal activity profiles for 14,000 human developmental craniofacial enhancers. We find that 56% of human craniofacial enhancers share chromatin accessibility in the mouse and we provide cell population- and embryonic stage-resolved predictions of their in vivo activity. Taken together, our data provide an expansive resource for genetic and developmental studies of human craniofacial development.
© 2024. The Author(s).
Conflict of interest statement
Bing Ren is a co-founder of Arima Genomics, Inc, and Epigenome Technologies, Inc. The remaining authors declare no competing interests.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
