

Enhancing prime editor activity by directed protein evolution in yeast
- PMID: 38453904
- PMCID: PMC10920827
- DOI: 10.1038/s41467-024-46107-z
Enhancing prime editor activity by directed protein evolution in yeast
Abstract
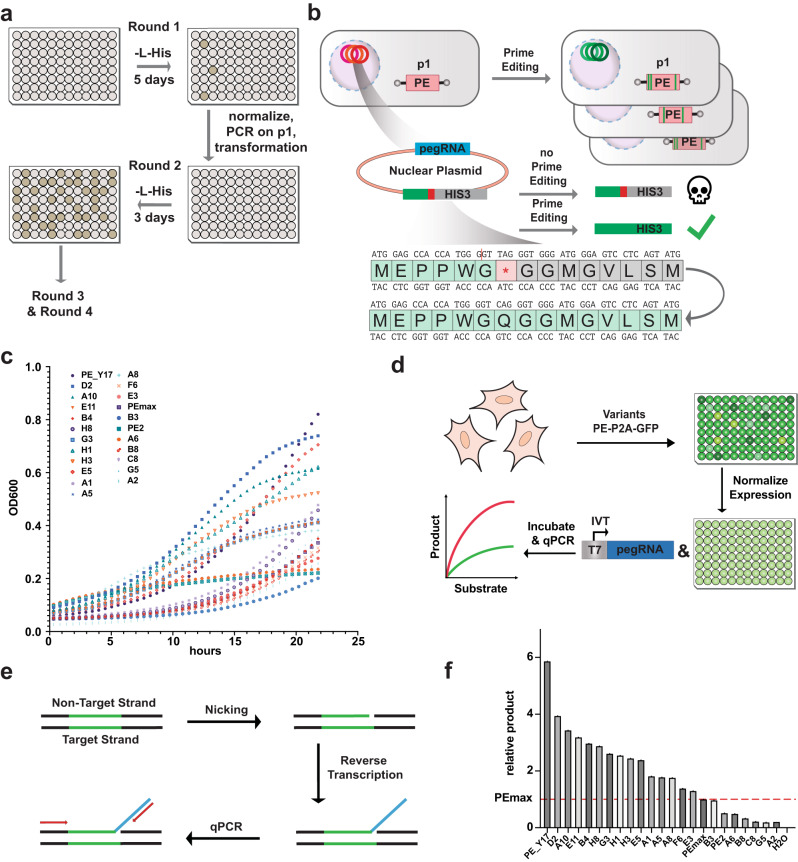
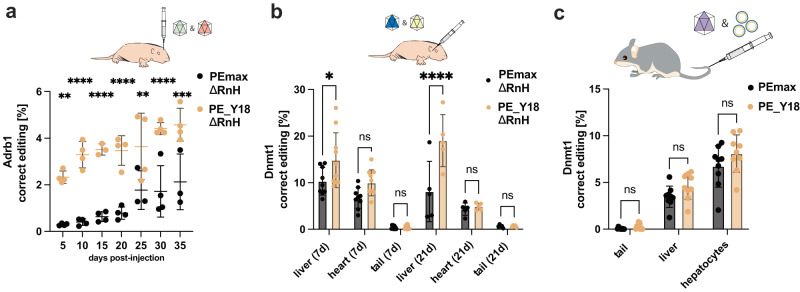
Prime editing is a highly versatile genome editing technology that enables the introduction of base substitutions, insertions, and deletions. However, compared to traditional Cas9 nucleases prime editors (PEs) are less active. In this study we use OrthoRep, a yeast-based platform for directed protein evolution, to enhance the editing efficiency of PEs. After several rounds of evolution with increased selection pressure, we identify multiple mutations that have a positive effect on PE activity in yeast cells and in biochemical assays. Combining the two most effective mutations - the A259D amino acid substitution in nCas9 and the K445T substitution in M-MLV RT - results in the variant PE_Y18. Delivery of PE_Y18, encoded on DNA, mRNA or as a ribonucleoprotein complex into mammalian cell lines increases editing rates up to 3.5-fold compared to PEmax. In addition, PE_Y18 supports higher prime editing rates when delivered in vivo into the liver or brain. Our study demonstrates proof-of-concept for the application of OrthoRep to optimize genome editing tools in eukaryotic cells.
© 2024. The Author(s).
Conflict of interest statement
Y.W. and G.S. have filed a patent application based on evolved PEmax variants as inventors (European Patent Application No. EP24152221.8). G.S. is a scientific advisor to Prime Medicine. The remaining authors declare no competing interests.
Figures


References
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
