

A guide to barley mutants
- PMID: 38454479
- PMCID: PMC10921644
- DOI: 10.1186/s41065-023-00304-w
A guide to barley mutants
Abstract
Background: Mutants have had a fundamental impact upon scientific and applied genetics. They have paved the way for the molecular and genomic era, and most of today's crop plants are derived from breeding programs involving mutagenic treatments.
Results: Barley (Hordeum vulgare L.) is one of the most widely grown cereals in the world and has a long history as a crop plant. Barley breeding started more than 100 years ago and large breeding programs have collected and generated a wide range of natural and induced mutants, which often were deposited in genebanks around the world. In recent years, an increased interest in genetic diversity has brought many historic mutants into focus because the collections are regarded as valuable resources for understanding the genetic control of barley biology and barley breeding. The increased interest has been fueled also by recent advances in genomic research, which provided new tools and possibilities to analyze and reveal the genetic diversity of mutant collections.
Conclusion: Since detailed knowledge about phenotypic characters of the mutants is the key to success of genetic and genomic studies, we here provide a comprehensive description of mostly morphological barley mutants. The review is closely linked to the International Database for Barley Genes and Barley Genetic Stocks ( bgs.nordgen.org ) where further details and additional images of each mutant described in this review can be found.
Keywords: Hordeum vulgare; Barley; Biodiversity; Cereal; Genebank; Induced mutants; Mutagenesis; Mutation; Triticeae.
© 2024. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures























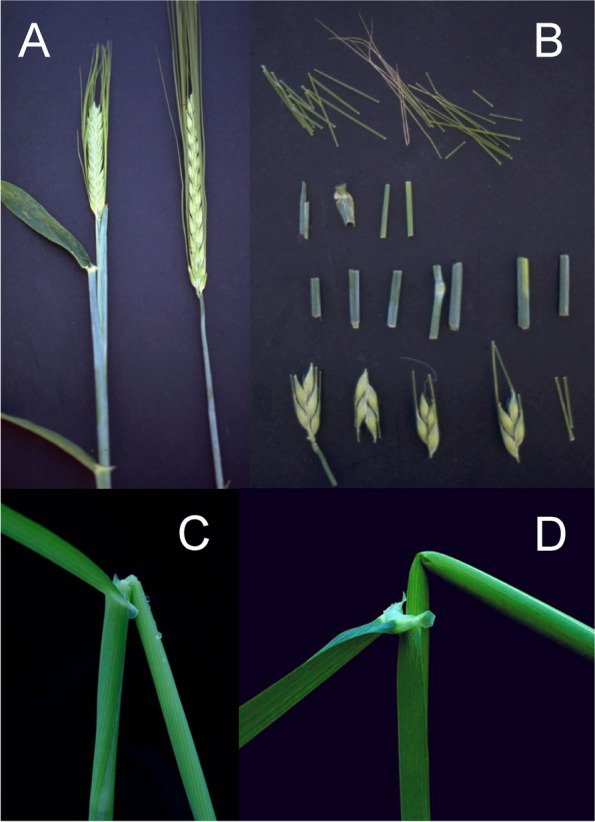



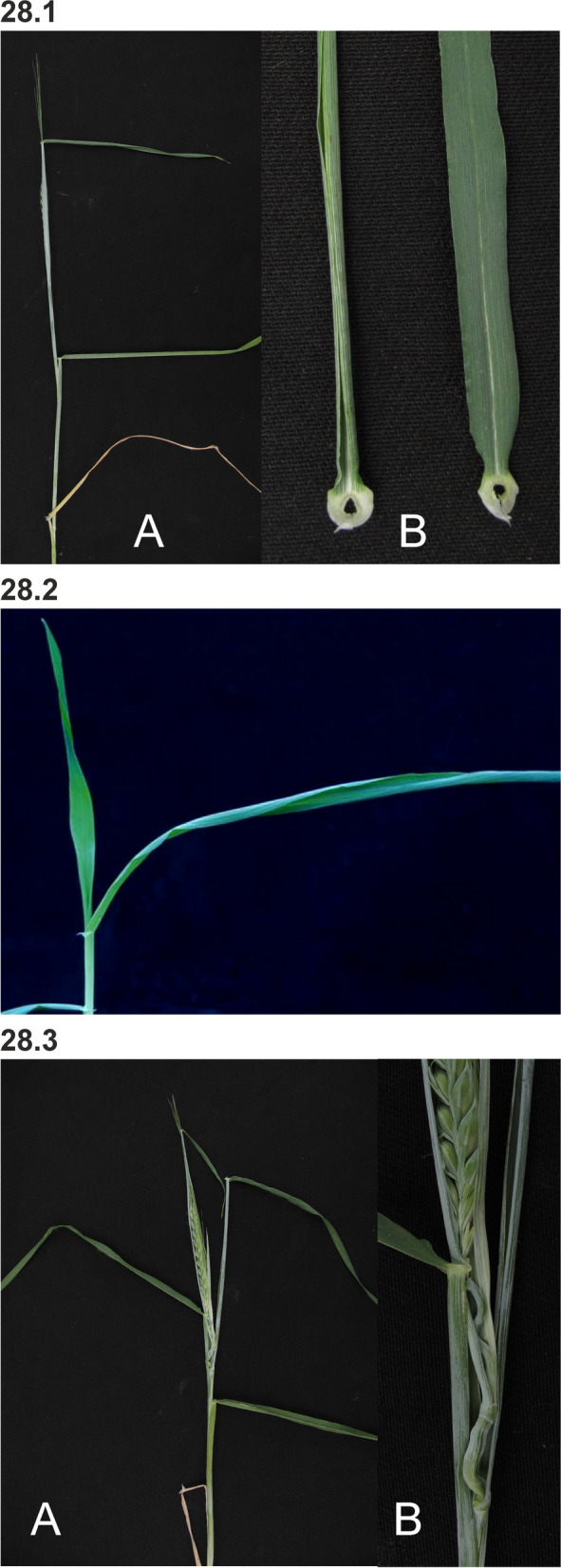










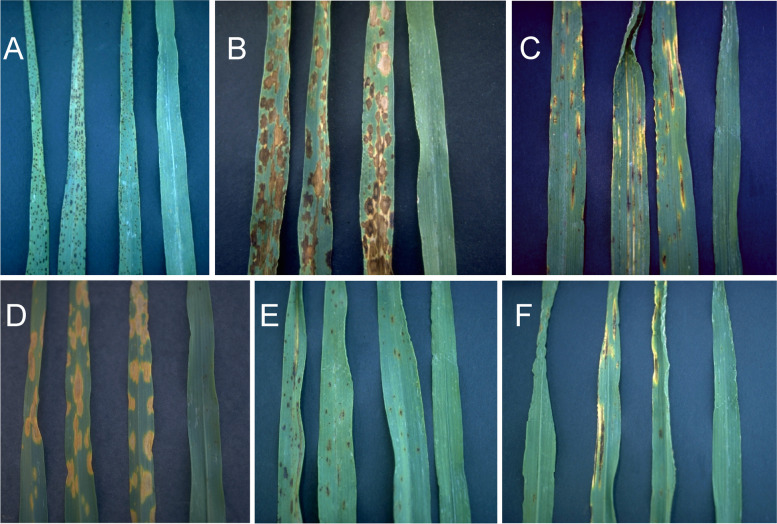

References
-
- Hansson M, Komatsuda T, Stein N, Muehlbauer GJ. Molecular mapping and cloning of genes and QTLs. In: Stein N, Muehlbauer GJ, editors. The barley genome. Cham, Switzerland: Springer Nature Switzerland AG; 2018. pp. 139–54.
-
- Gustafsson Å. Studies on the genetic basis of chlorophyll formulation and the machanism of induced mutatingy. Hereditas. 1938;24:33–93.
-
- Gustafsson Å. The mutation system of the chlorophyll apparatus. Lunds Universitets Årsskrift N F Avd. 1940;2(36):1–40.
-
- Hallqvist C. Chlorophyllmutanten bei Gerste. Hereditas. 1924;5:49–83. doi: 10.1111/j.1601-5223.1924.tb03123.x. - DOI
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
