

p66Shc signaling and autophagy impact on C2C12 myoblast differentiation during senescence
- PMID: 38459002
- PMCID: PMC10923948
- DOI: 10.1038/s41419-024-06582-0
p66Shc signaling and autophagy impact on C2C12 myoblast differentiation during senescence
Abstract
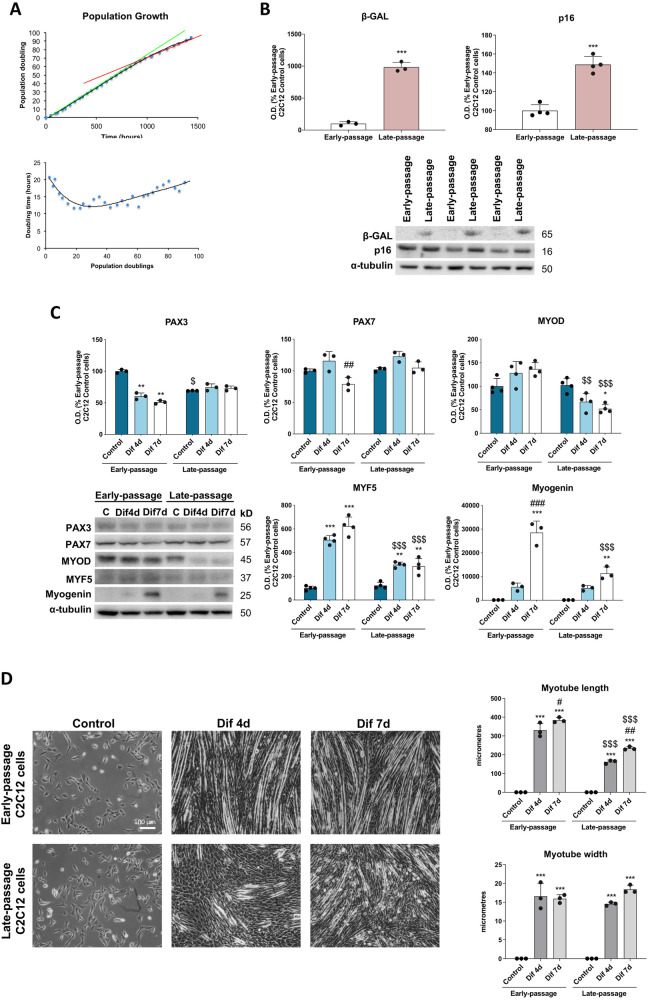
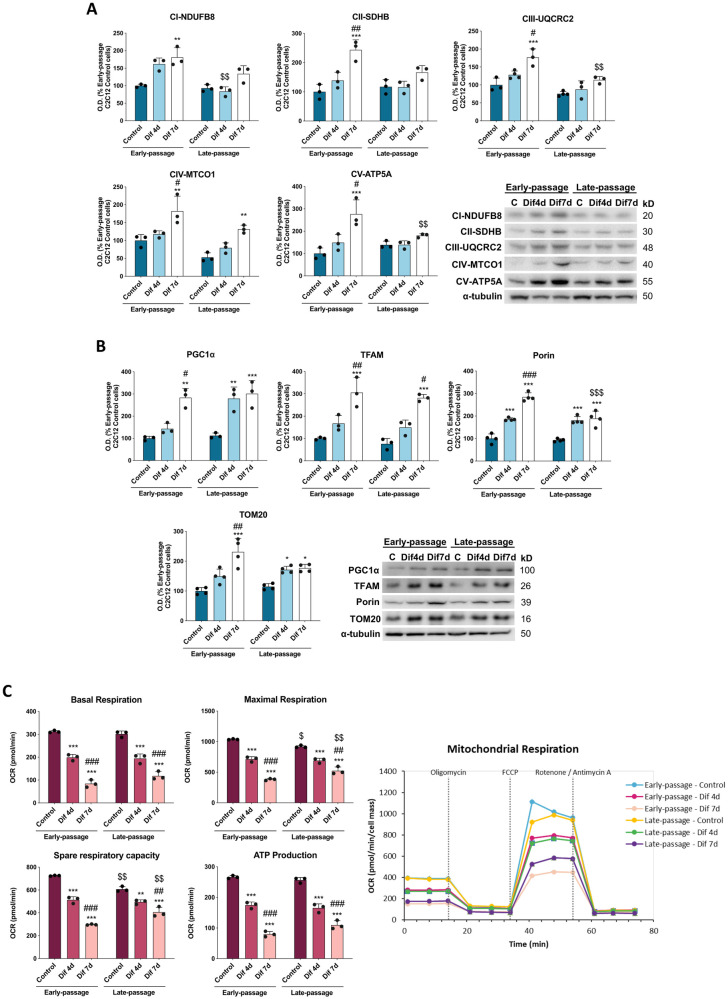
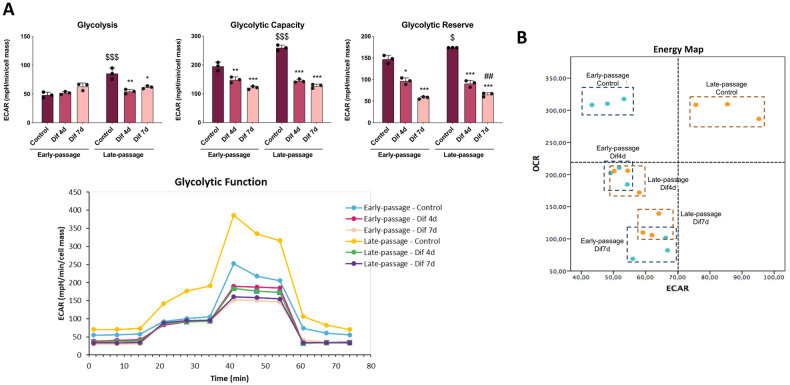
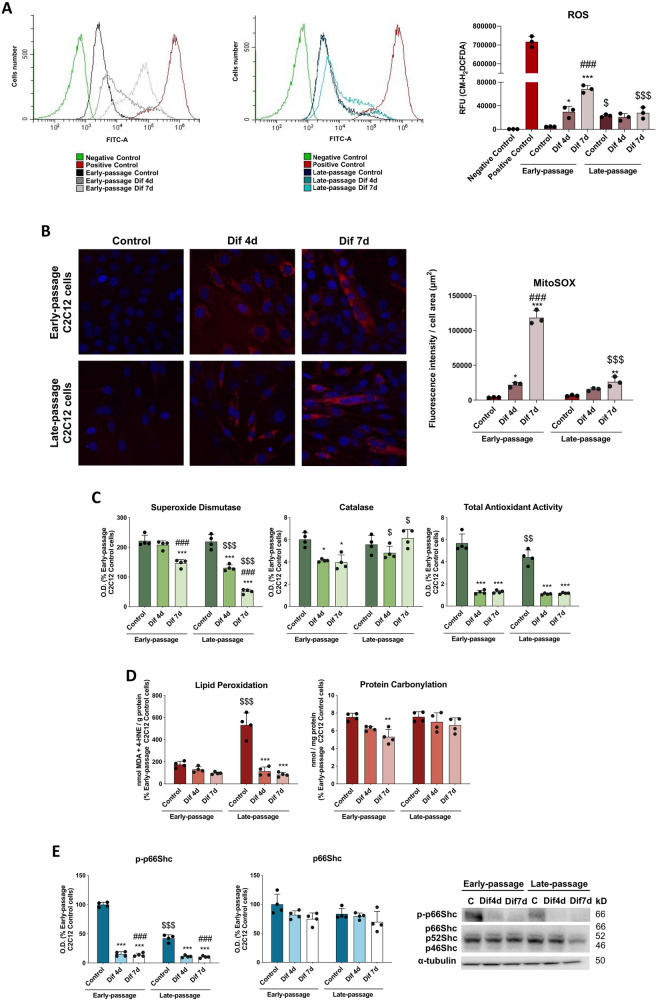
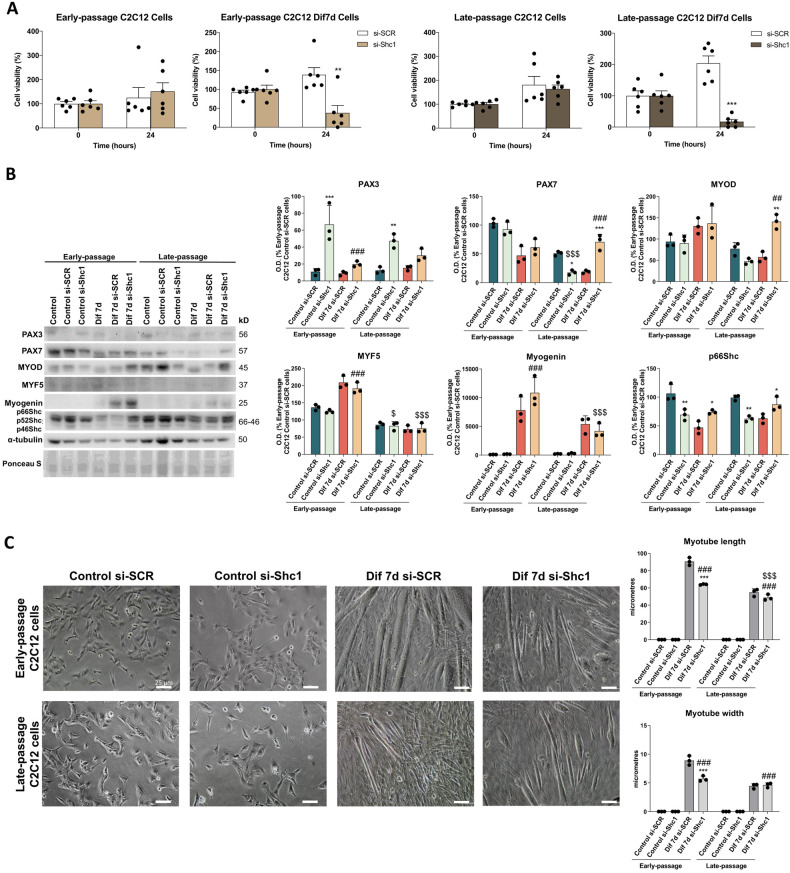
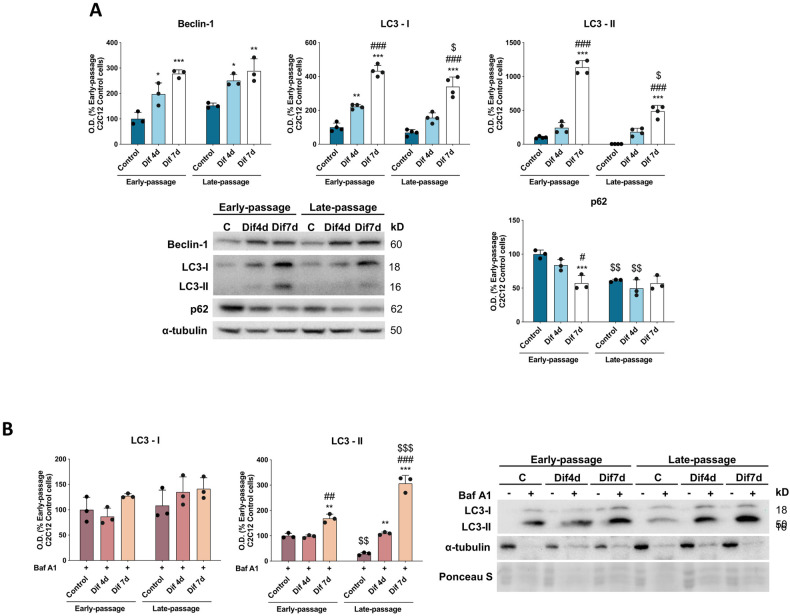
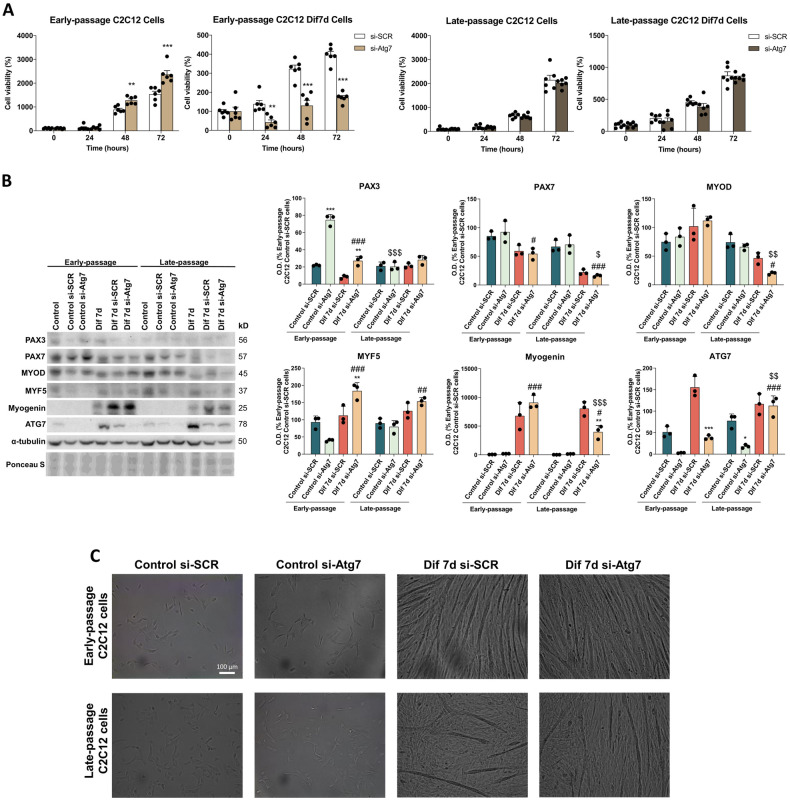
During aging, muscle regenerative capacities decline, which is concomitant with the loss of satellite cells that enter in a state of irreversible senescence. However, what mechanisms are involved in myogenic senescence and differentiation are largely unknown. Here, we showed that early-passage or "young" C2C12 myoblasts activated the redox-sensitive p66Shc signaling pathway, exhibited a strong antioxidant protection and a bioenergetic profile relying predominantly on OXPHOS, responses that decrease progressively during differentiation. Furthermore, autophagy was increased in myotubes. Otherwise, late-passage or "senescent" myoblasts led to a highly metabolic profile, relying on both OXPHOS and glycolysis, that may be influenced by the loss of SQSTM1/p62 which tightly regulates the metabolic shift from aerobic glycolysis to OXPHOS. Furthermore, during differentiation of late-passage C2C12 cells, both p66Shc signaling and autophagy were impaired and this coincides with reduced myogenic capacity. Our findings recognized that the lack of p66Shc compromises the proliferation and the onset of the differentiation of C2C12 myoblasts. Moreover, the Atg7 silencing favored myoblasts growth, whereas interfered in the viability of differentiated myotubes. Then, our work demonstrates that the p66Shc signaling pathway, which highly influences cellular metabolic status and oxidative environment, is critical for the myogenic commitment and differentiation of C2C12 cells. Our findings also support that autophagy is essential for the metabolic switch observed during the differentiation of C2C12 myoblasts, confirming how its regulation determines cell fate. The regulatory roles of p66Shc and autophagy mechanisms on myogenesis require future attention as possible tools that could predict and measure the aging-related state of frailty and disability.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
