

PMI-controlled mannose metabolism and glycosylation determines tissue tolerance and virus fitness
- PMID: 38459021
- PMCID: PMC10923791
- DOI: 10.1038/s41467-024-46415-4
PMI-controlled mannose metabolism and glycosylation determines tissue tolerance and virus fitness
Abstract
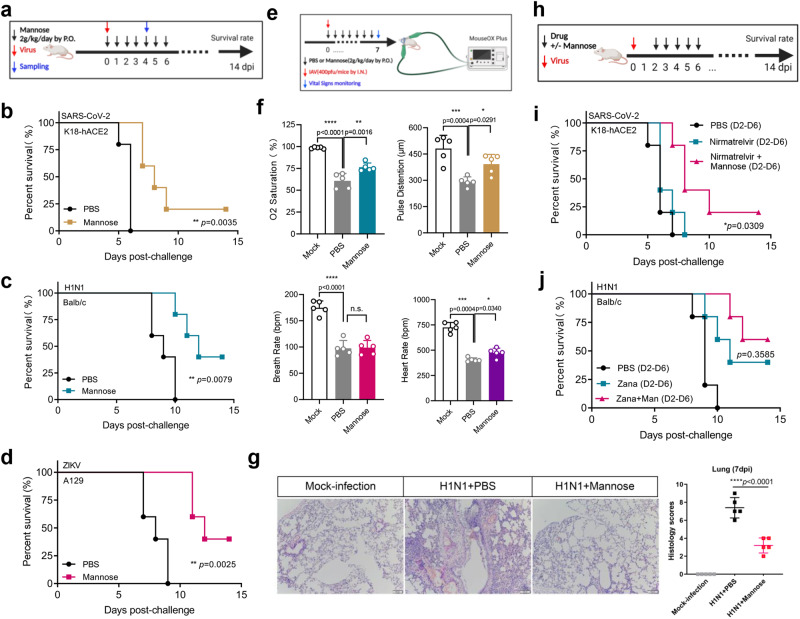
Host survival depends on the elimination of virus and mitigation of tissue damage. Herein, we report the modulation of D-mannose flux rewires the virus-triggered immunometabolic response cascade and reduces tissue damage. Safe and inexpensive D-mannose can compete with glucose for the same transporter and hexokinase. Such competitions suppress glycolysis, reduce mitochondrial reactive-oxygen-species and succinate-mediated hypoxia-inducible factor-1α, and thus reduce virus-induced proinflammatory cytokine production. The combinatorial treatment by D-mannose and antiviral monotherapy exhibits in vivo synergy despite delayed antiviral treatment in mouse model of virus infections. Phosphomannose isomerase (PMI) knockout cells are viable, whereas addition of D-mannose to the PMI knockout cells blocks cell proliferation, indicating that PMI activity determines the beneficial effect of D-mannose. PMI inhibition suppress a panel of virus replication via affecting host and viral surface protein glycosylation. However, D-mannose does not suppress PMI activity or virus fitness. Taken together, PMI-centered therapeutic strategy clears virus infection while D-mannose treatment reprograms glycolysis for control of collateral damage.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures



Similar articles
-
Mannose and PMI depletion overcomes radiation resistance in HPV-negative head and neck cancer.Cell Commun Signal. 2025 Apr 21;23(1):189. doi: 10.1186/s12964-025-02204-0. Cell Commun Signal. 2025. PMID: 40259370 Free PMC article.
-
The relative contribution of mannose salvage pathways to glycosylation in PMI-deficient mouse embryonic fibroblast cells.FEBS J. 2008 Feb;275(4):788-98. doi: 10.1111/j.1742-4658.2008.06246.x. Epub 2008 Jan 22. FEBS J. 2008. PMID: 18215164
-
Mannose impairs tumour growth and enhances chemotherapy.Nature. 2018 Nov;563(7733):719-723. doi: 10.1038/s41586-018-0729-3. Epub 2018 Nov 21. Nature. 2018. PMID: 30464341
-
Mannose metabolism: more than meets the eye.Biochem Biophys Res Commun. 2014 Oct 17;453(2):220-8. doi: 10.1016/j.bbrc.2014.06.021. Epub 2014 Jun 12. Biochem Biophys Res Commun. 2014. PMID: 24931670 Free PMC article. Review.
-
The Antiviral Effects of 2-Deoxy-D-glucose (2-DG), a Dual D-Glucose and D-Mannose Mimetic, against SARS-CoV-2 and Other Highly Pathogenic Viruses.Molecules. 2022 Sep 12;27(18):5928. doi: 10.3390/molecules27185928. Molecules. 2022. PMID: 36144664 Free PMC article. Review.
Cited by
-
Mannose Promotes β-Amyloid Pathology by Regulating BACE1 Glycosylation in Alzheimer's Disease.Adv Sci (Weinh). 2025 Mar;12(9):e2409105. doi: 10.1002/advs.202409105. Epub 2025 Jan 14. Adv Sci (Weinh). 2025. PMID: 39807036 Free PMC article.
-
D-mannose promotes diabetic wound healing through inhibiting advanced glycation end products formation in keratinocytes.Mol Med. 2025 Jan 18;31(1):15. doi: 10.1186/s10020-025-01070-3. Mol Med. 2025. PMID: 39827347 Free PMC article.
-
Classical swine fever virus utilizes stearoyl-CoA desaturase 1-mediated lipid metabolism to facilitate viral replication.J Virol. 2025 Jun 17;99(6):e0055125. doi: 10.1128/jvi.00551-25. Epub 2025 May 19. J Virol. 2025. PMID: 40387378 Free PMC article.
-
Mannose and PMI depletion overcomes radiation resistance in HPV-negative head and neck cancer.Cell Commun Signal. 2025 Apr 21;23(1):189. doi: 10.1186/s12964-025-02204-0. Cell Commun Signal. 2025. PMID: 40259370 Free PMC article.
-
Sugar Functionalized Collagen Material for Local Modulation of Innate Immunity.Adv Sci (Weinh). 2025 Aug;12(31):e2415364. doi: 10.1002/advs.202415364. Epub 2025 May 24. Adv Sci (Weinh). 2025. PMID: 40411409 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
