

Learning structural heterogeneity from cryo-electron sub-tomograms with tomoDRGN
- PMID: 38459385
- PMCID: PMC11655136
- DOI: 10.1038/s41592-024-02210-z
Learning structural heterogeneity from cryo-electron sub-tomograms with tomoDRGN
Abstract
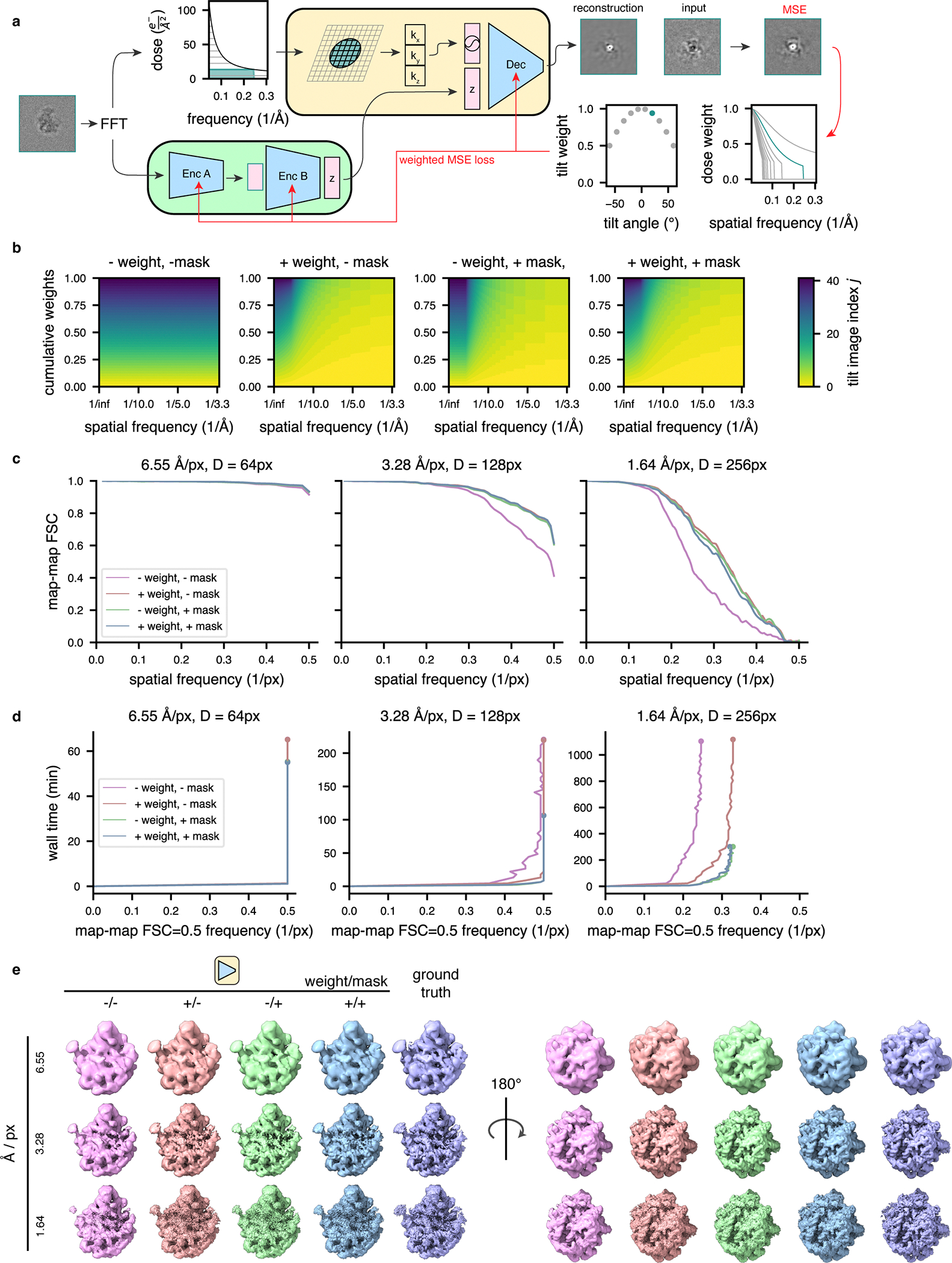
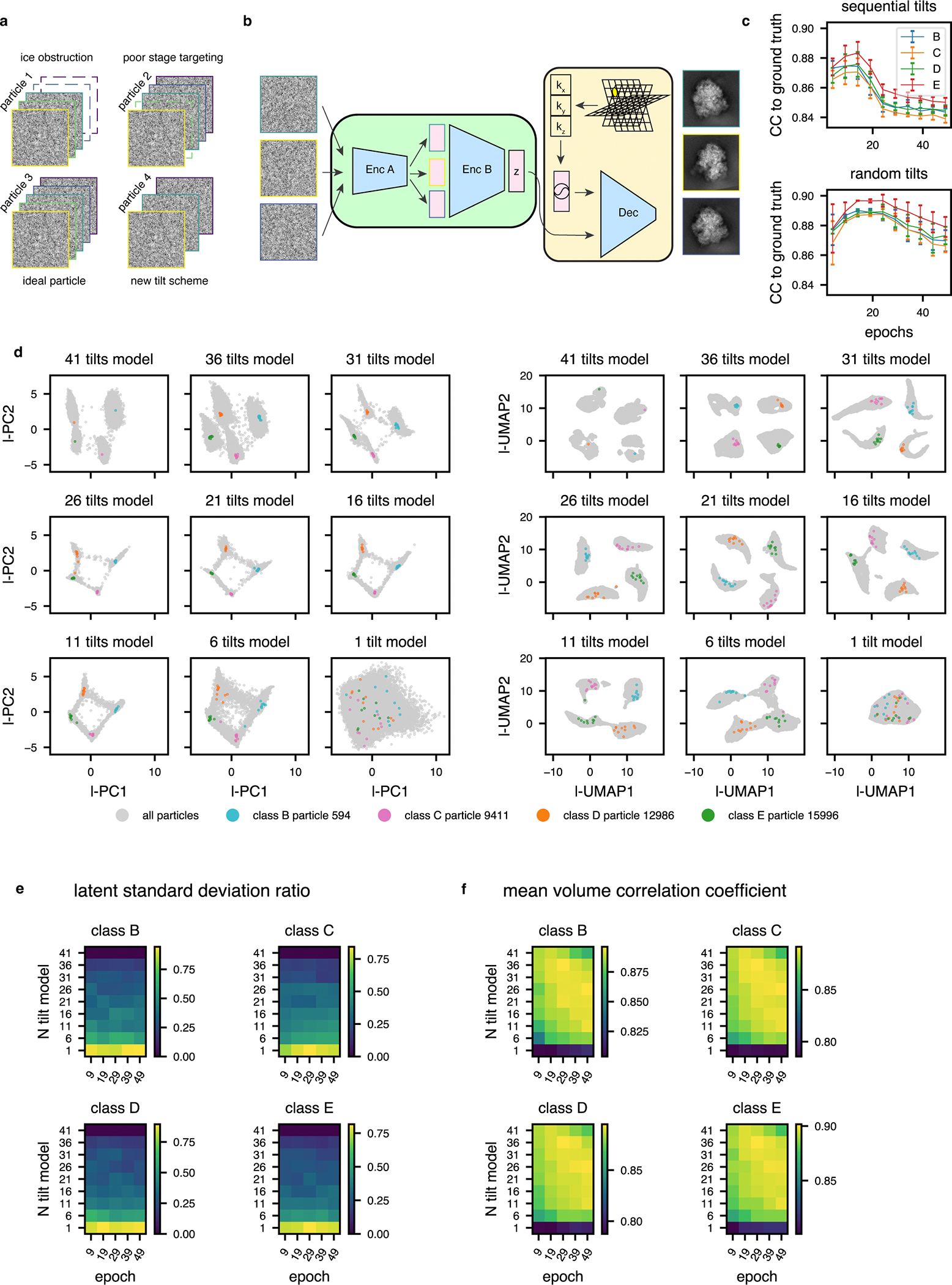
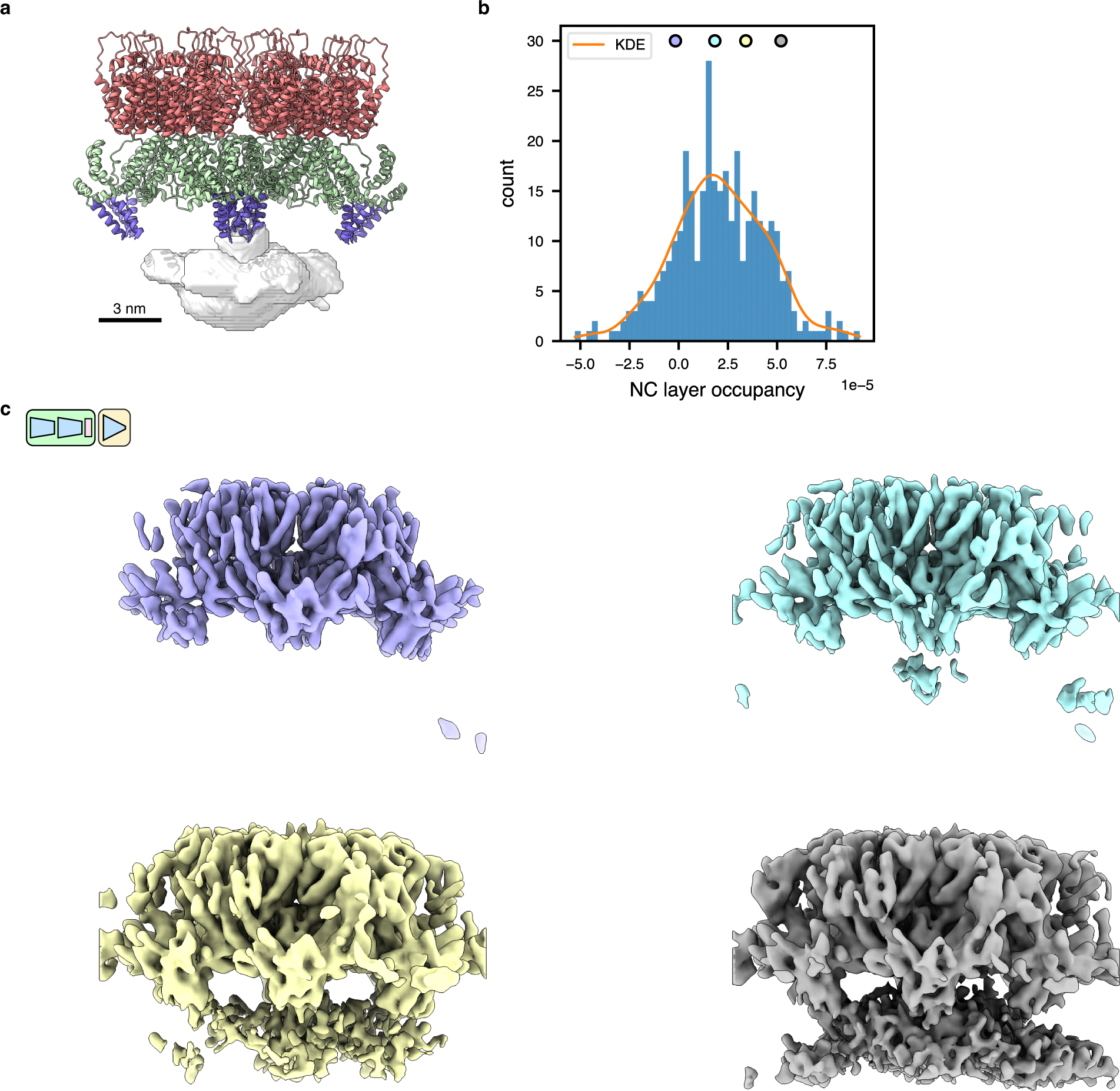
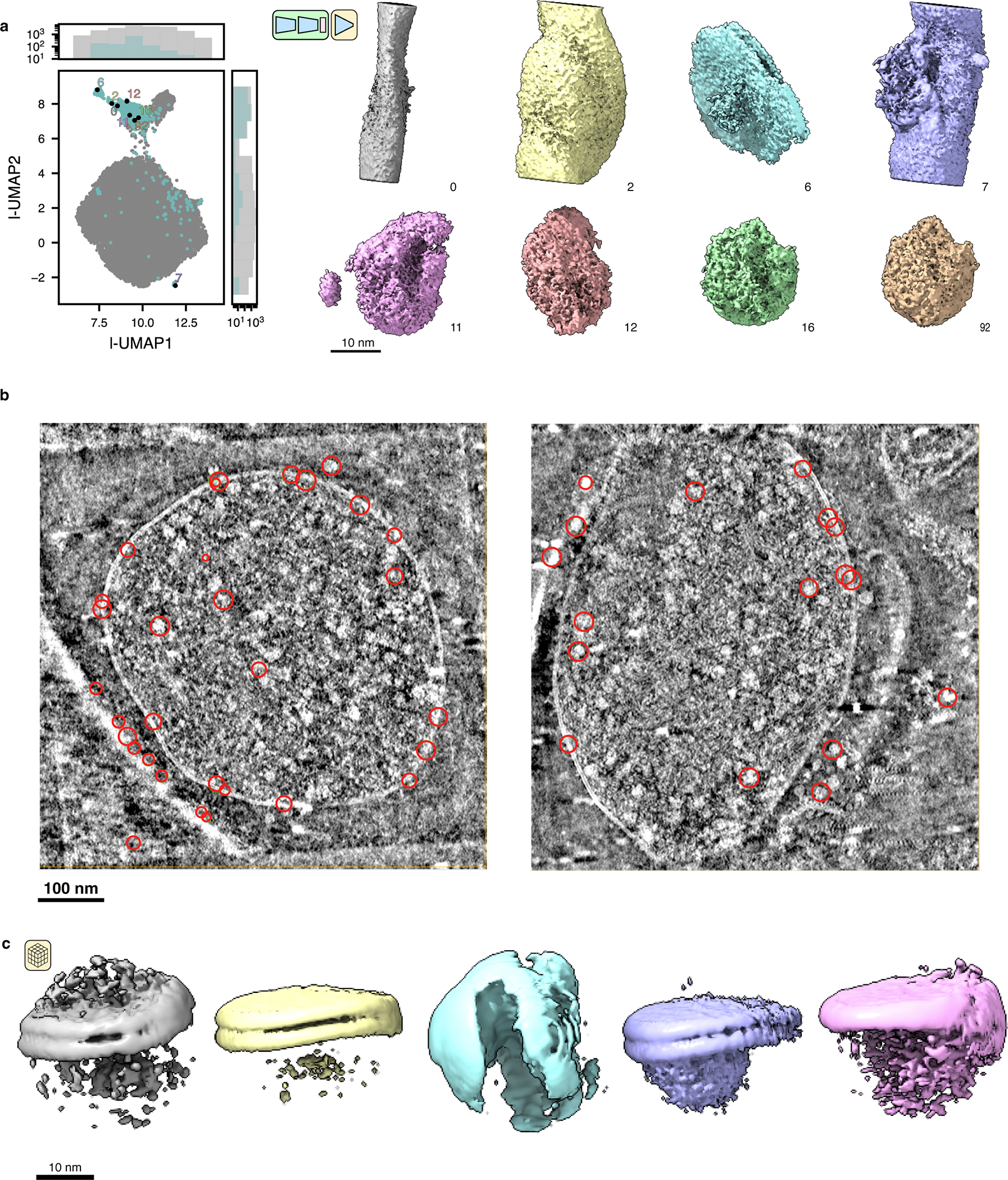
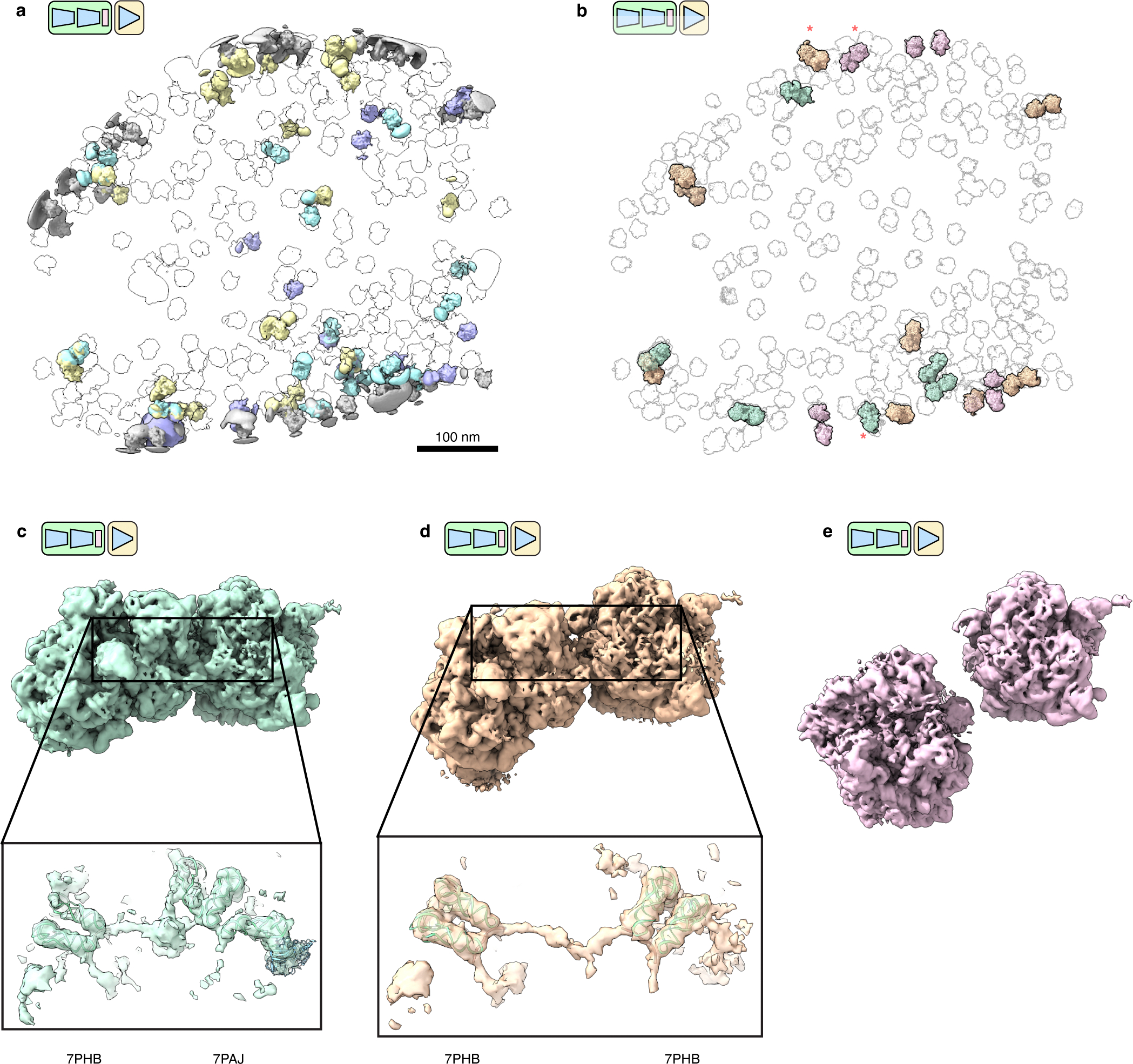
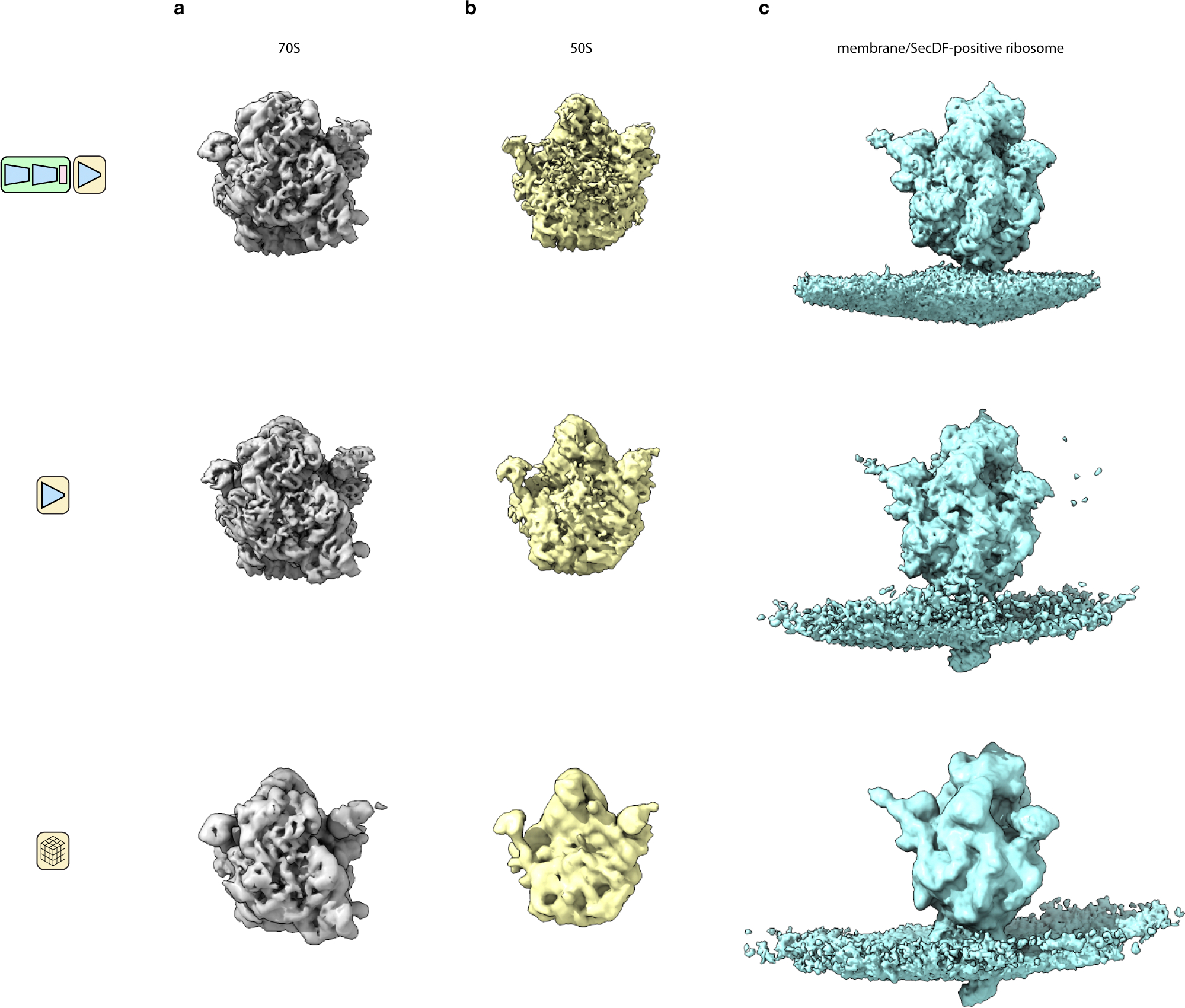
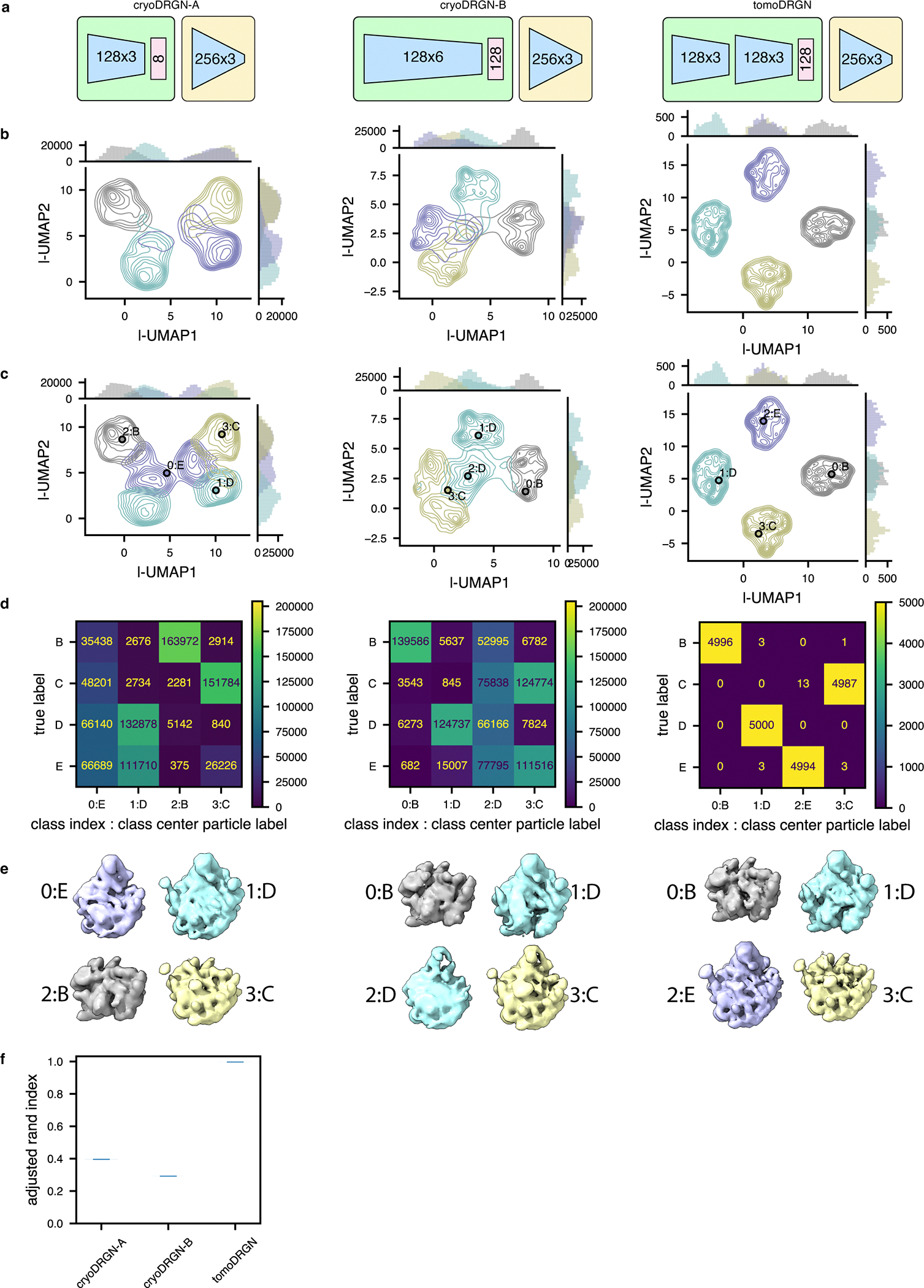
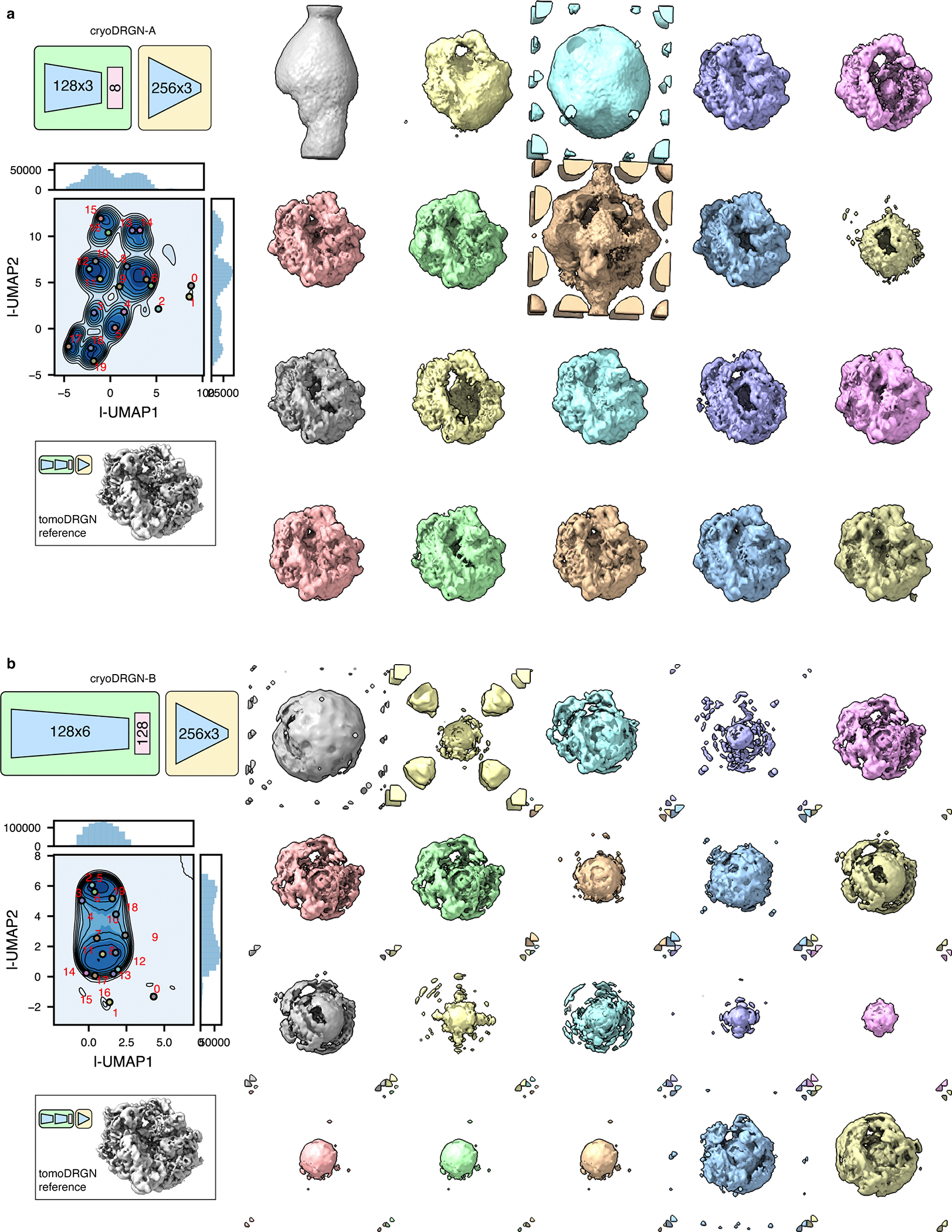
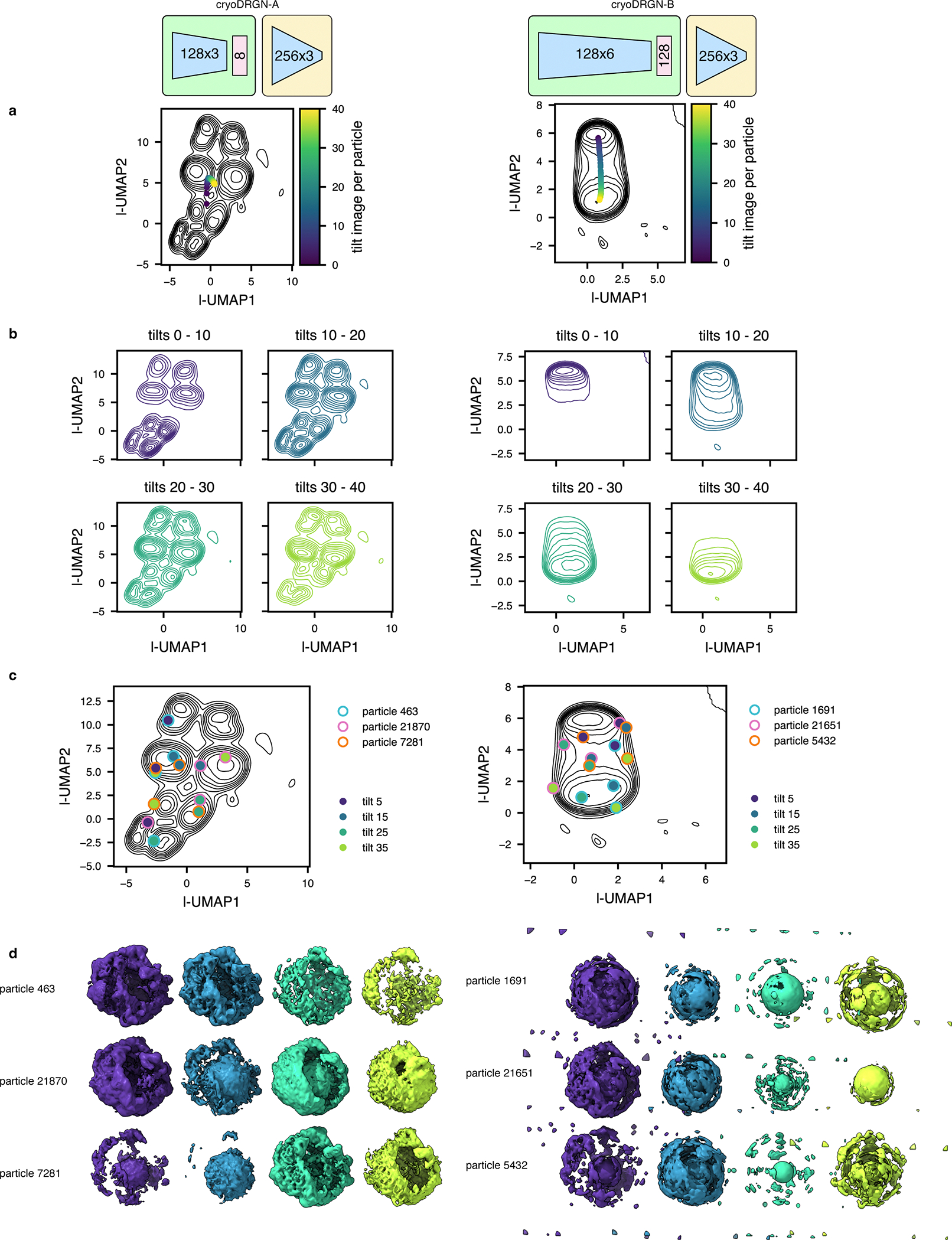
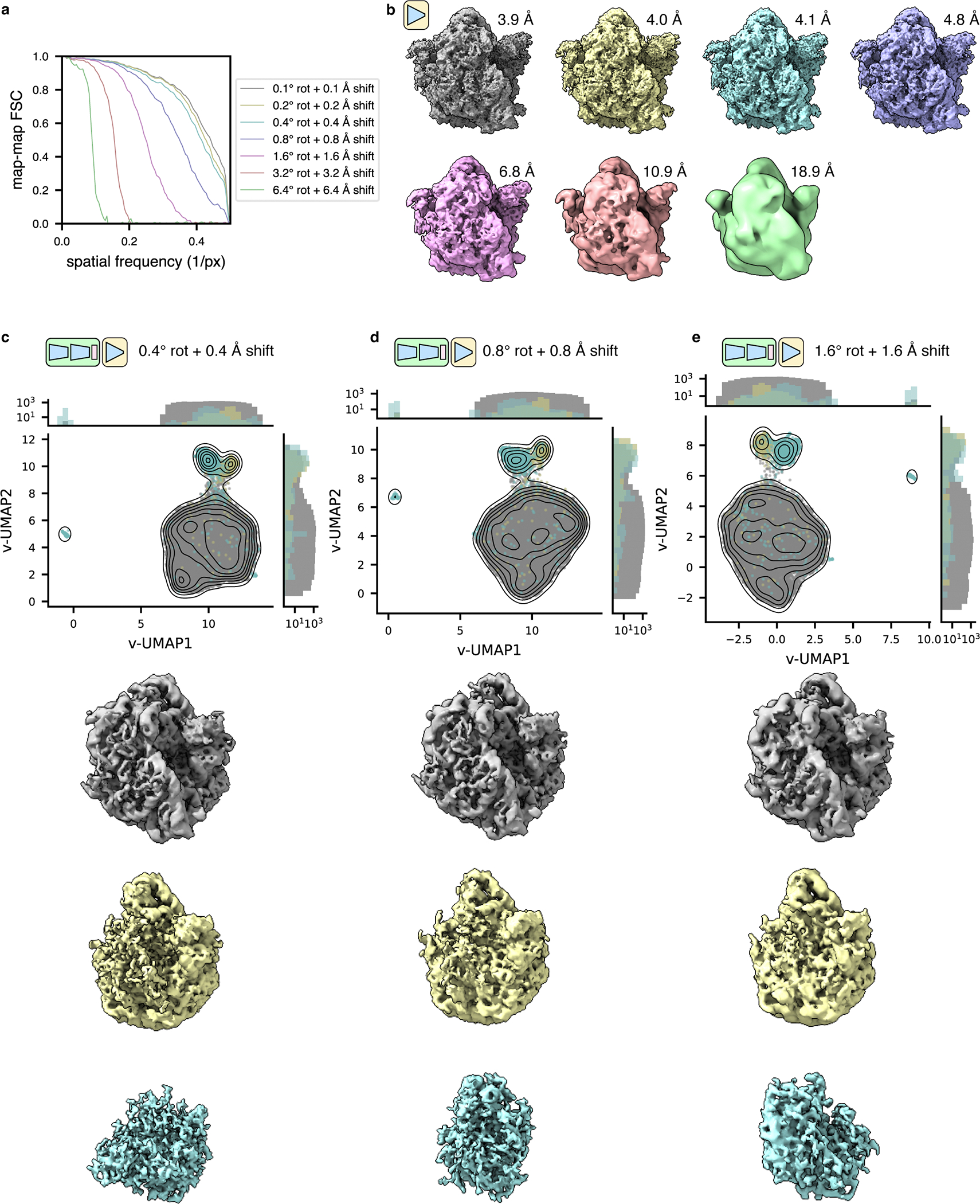
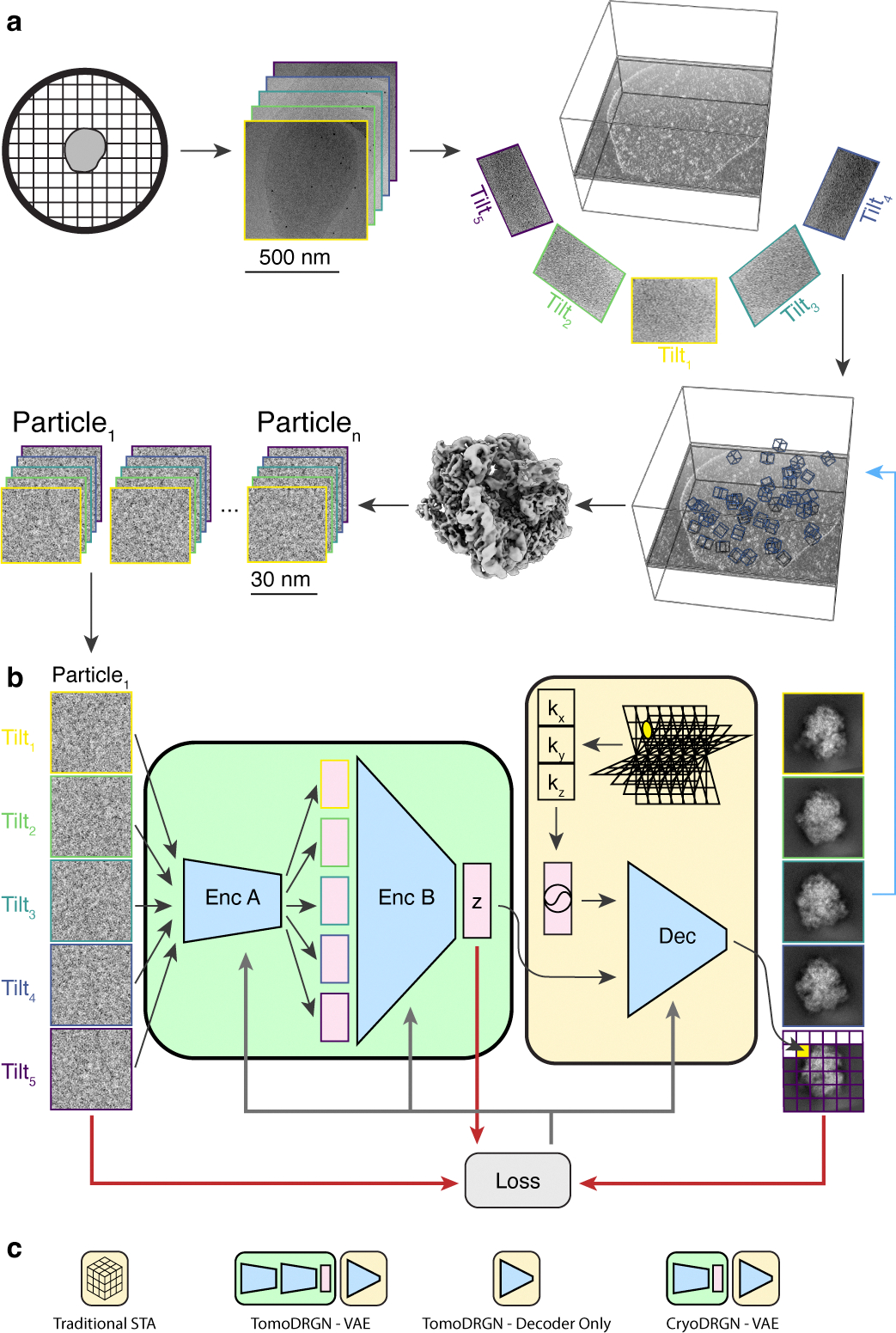
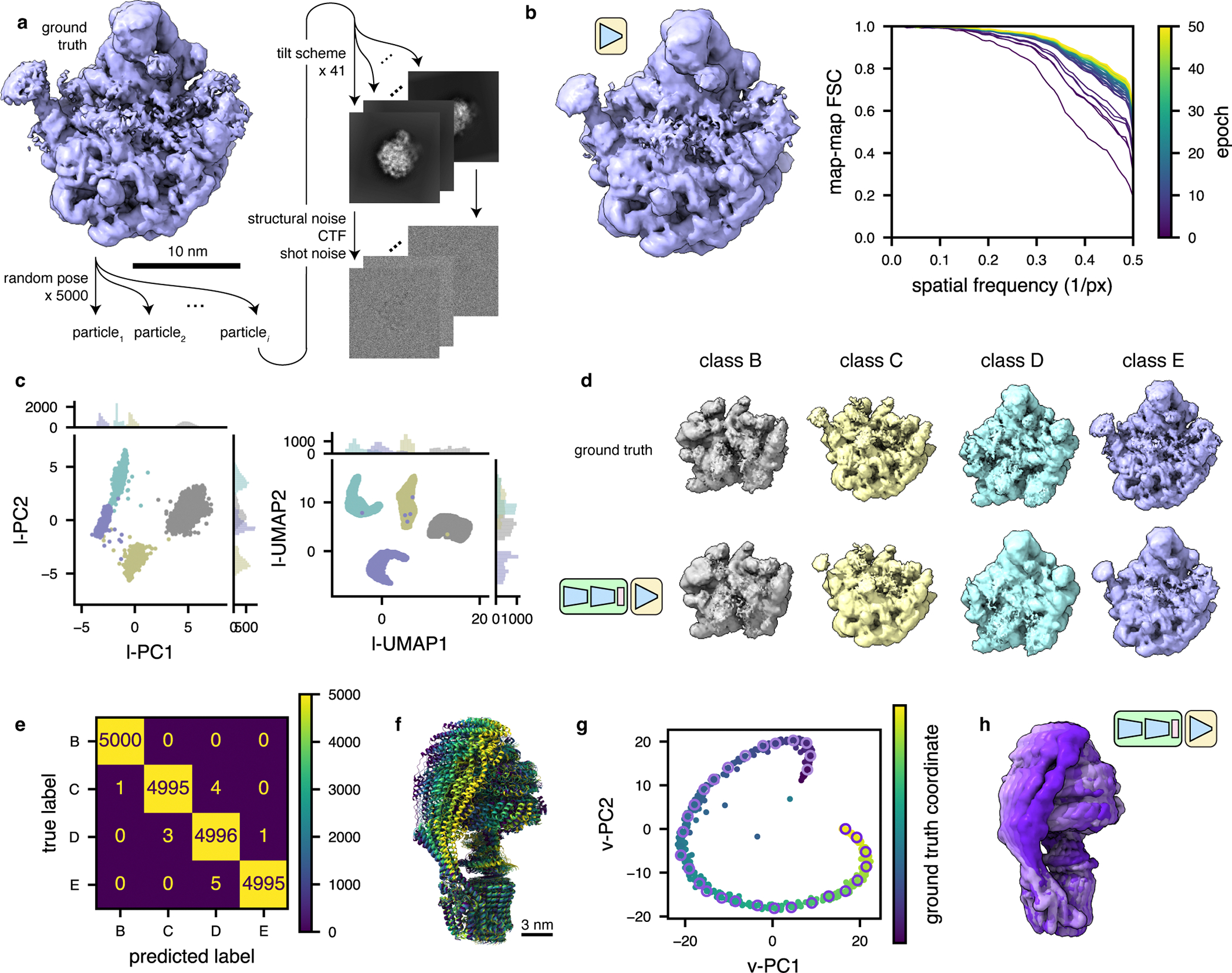
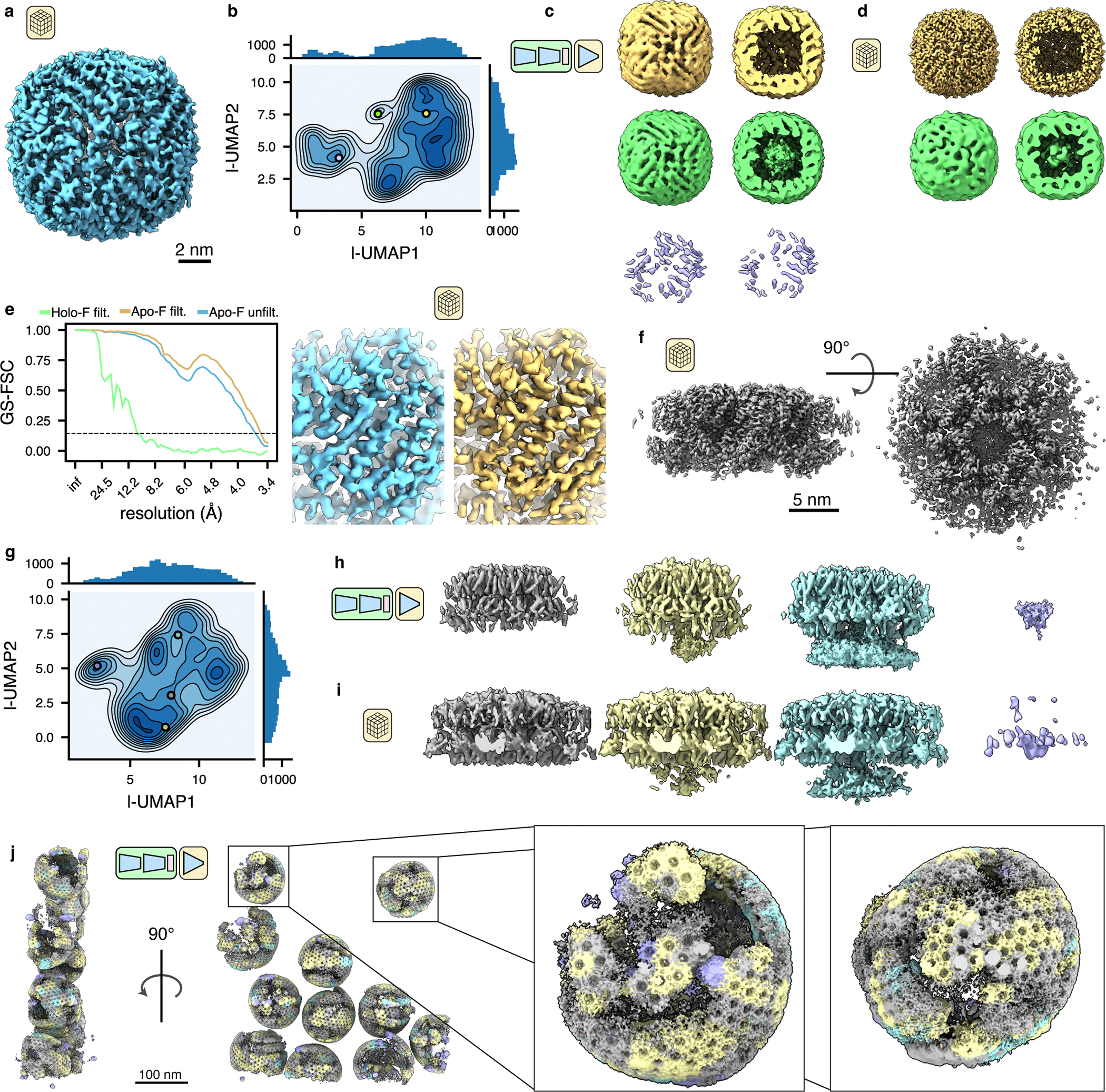
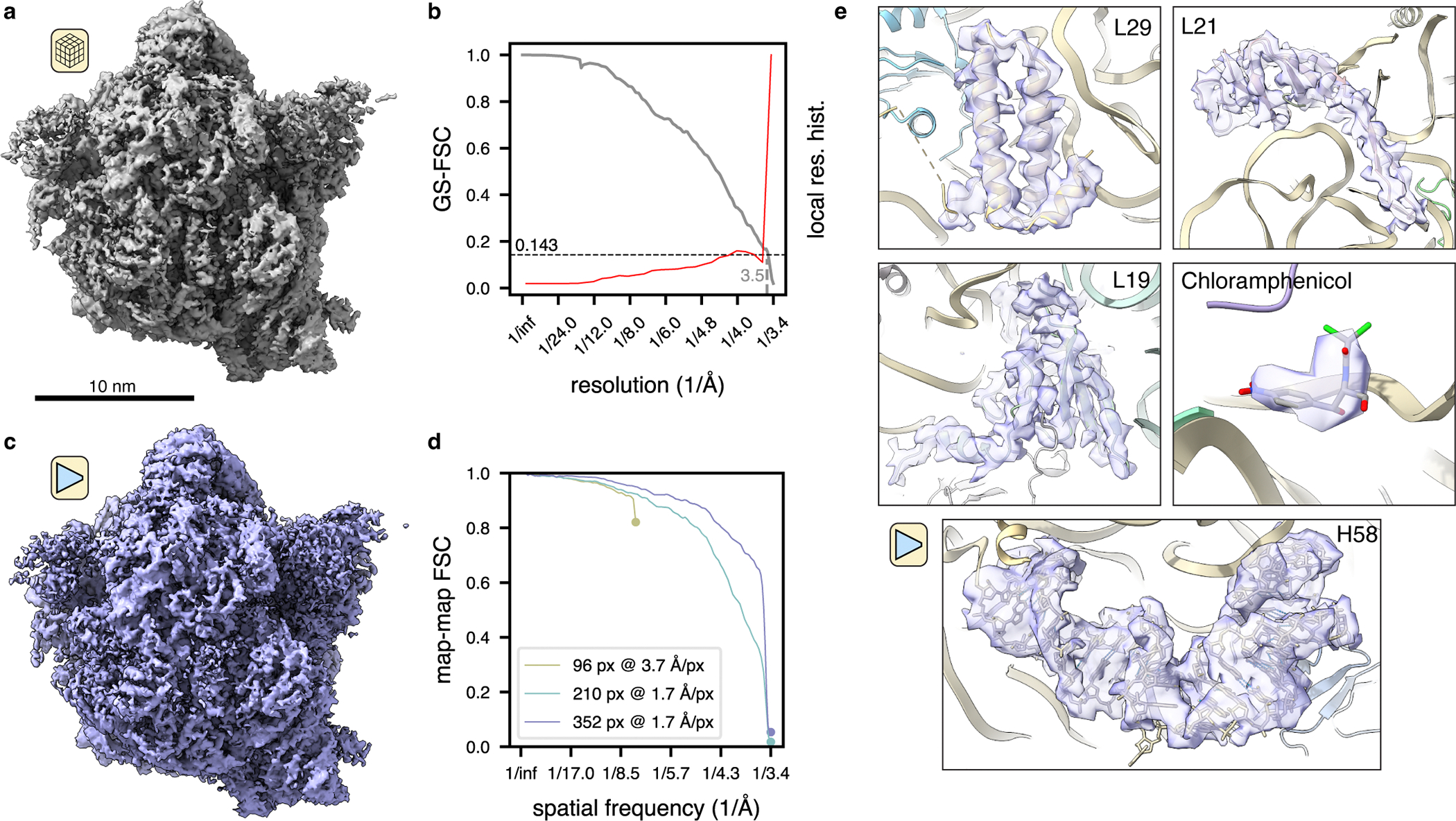
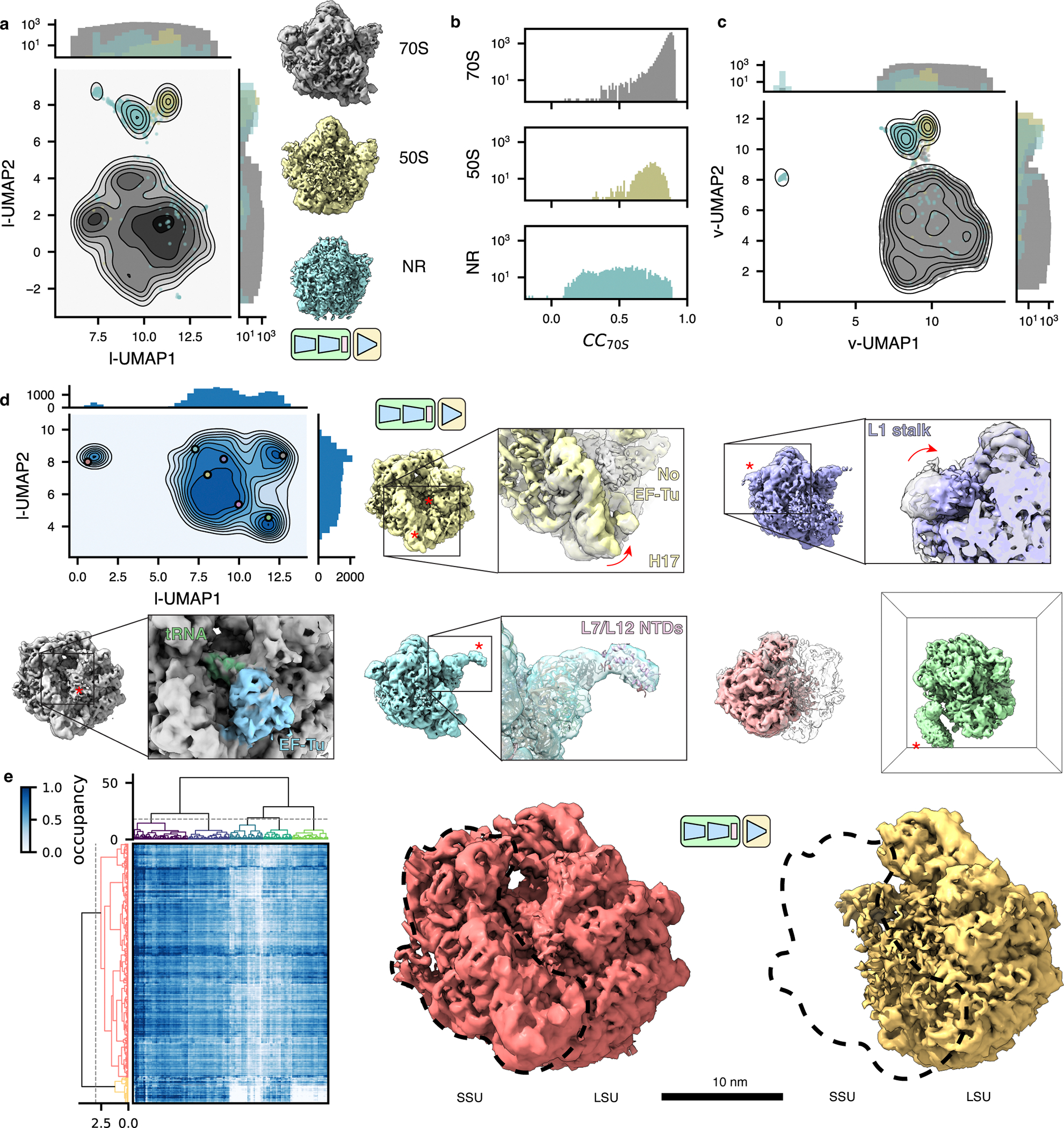
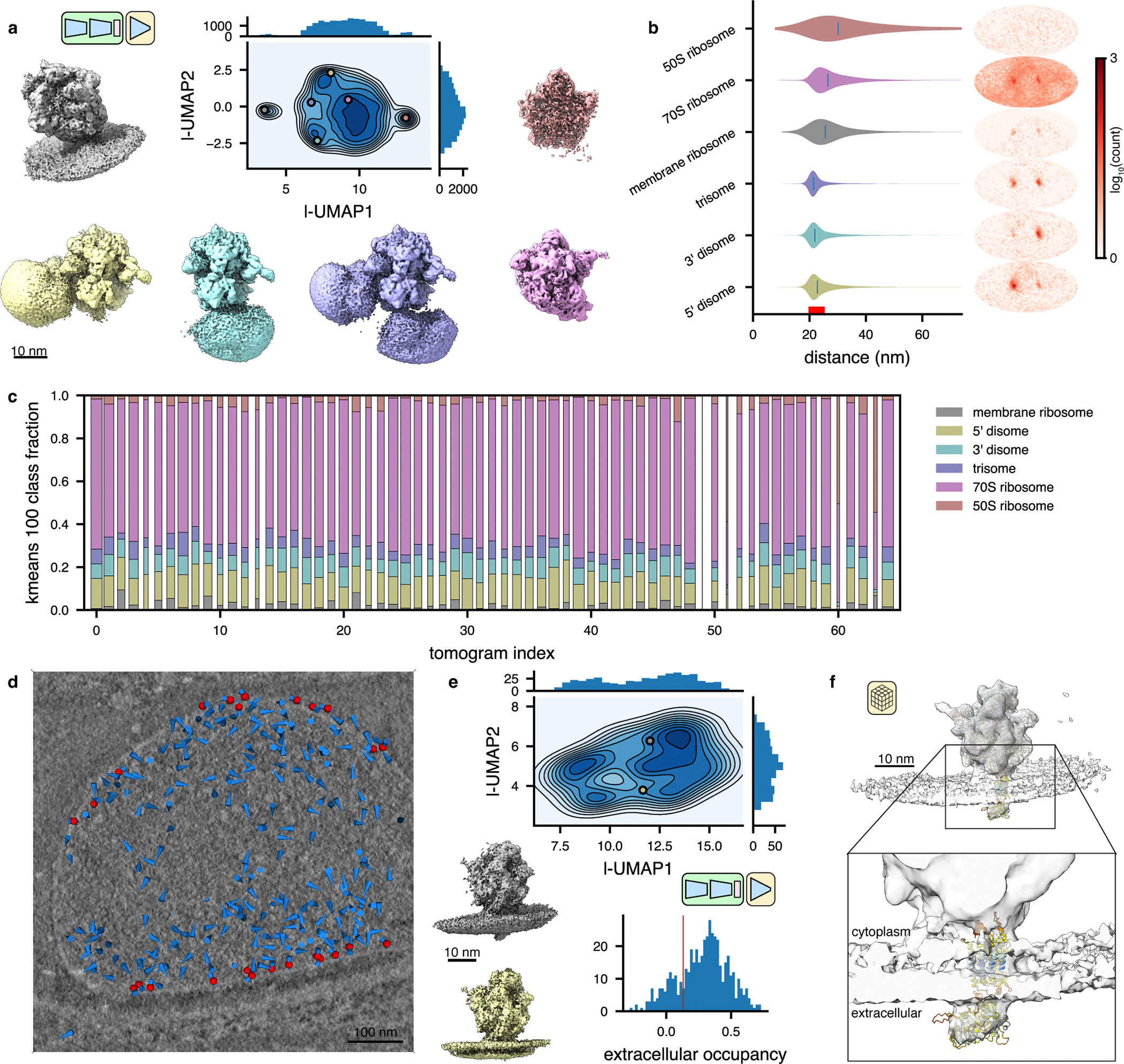
Cryo-electron tomography (cryo-ET) enables observation of macromolecular complexes in their native, spatially contextualized cellular environment. Cryo-ET processing software to visualize such complexes at nanometer resolution via iterative alignment and averaging are well developed but rely upon assumptions of structural homogeneity among the complexes of interest. Recently developed tools allow for some assessment of structural diversity but have limited capacity to represent highly heterogeneous structures, including those undergoing continuous conformational changes. Here we extend the highly expressive cryoDRGN (Deep Reconstructing Generative Networks) deep learning architecture, originally created for single-particle cryo-electron microscopy analysis, to cryo-ET. Our new tool, tomoDRGN, learns a continuous low-dimensional representation of structural heterogeneity in cryo-ET datasets while also learning to reconstruct heterogeneous structural ensembles supported by the underlying data. Using simulated and experimental data, we describe and benchmark architectural choices within tomoDRGN that are uniquely necessitated and enabled by cryo-ET. We additionally illustrate tomoDRGN's efficacy in analyzing diverse datasets, using it to reveal high-level organization of human immunodeficiency virus (HIV) capsid complexes assembled in virus-like particles and to resolve extensive structural heterogeneity among ribosomes imaged in situ.
© 2024. The Author(s), under exclusive licence to Springer Nature America, Inc.
Conflict of interest statement
COMPETING INTERESTS STATEMENT
The authors declare no competing interests.
Figures
Update of
-
Learning structural heterogeneity from cryo-electron sub-tomograms with tomoDRGN.bioRxiv [Preprint]. 2023 Jun 2:2023.05.31.542975. doi: 10.1101/2023.05.31.542975. bioRxiv. 2023. Update in: Nat Methods. 2024 Aug;21(8):1525-1536. doi: 10.1038/s41592-024-02210-z. PMID: 37398315 Free PMC article. Updated. Preprint.
Similar articles
-
Learning structural heterogeneity from cryo-electron sub-tomograms with tomoDRGN.bioRxiv [Preprint]. 2023 Jun 2:2023.05.31.542975. doi: 10.1101/2023.05.31.542975. bioRxiv. 2023. Update in: Nat Methods. 2024 Aug;21(8):1525-1536. doi: 10.1038/s41592-024-02210-z. PMID: 37398315 Free PMC article. Updated. Preprint.
-
CryoDRGN-ET: deep reconstructing generative networks for visualizing dynamic biomolecules inside cells.Nat Methods. 2024 Aug;21(8):1537-1545. doi: 10.1038/s41592-024-02340-4. Epub 2024 Jul 18. Nat Methods. 2024. PMID: 39025970
-
Current data processing strategies for cryo-electron tomography and subtomogram averaging.Biochem J. 2021 May 28;478(10):1827-1845. doi: 10.1042/BCJ20200715. Biochem J. 2021. PMID: 34003255 Free PMC article. Review.
-
Using Tomoauto: A Protocol for High-throughput Automated Cryo-electron Tomography.J Vis Exp. 2016 Jan 30;(107):e53608. doi: 10.3791/53608. J Vis Exp. 2016. PMID: 26863591 Free PMC article.
-
Subtomogram averaging from cryo-electron tomograms.Methods Cell Biol. 2019;152:217-259. doi: 10.1016/bs.mcb.2019.04.003. Epub 2019 May 15. Methods Cell Biol. 2019. PMID: 31326022 Review.
Cited by
-
Rendering protein structures inside cells at the atomic level with Unreal Engine.bioRxiv [Preprint]. 2024 Mar 9:2023.12.08.570879. doi: 10.1101/2023.12.08.570879. bioRxiv. 2024. PMID: 38496473 Free PMC article. Preprint.
-
Bridging structural and cell biology with cryo-electron microscopy.Nature. 2024 Apr;628(8006):47-56. doi: 10.1038/s41586-024-07198-2. Epub 2024 Apr 3. Nature. 2024. PMID: 38570716 Free PMC article. Review.
-
A new age in structural S-layer biology: Experimental and in silico milestones.J Biol Chem. 2025 Jun;301(6):110205. doi: 10.1016/j.jbc.2025.110205. Epub 2025 May 8. J Biol Chem. 2025. PMID: 40345586 Free PMC article. Review.
-
In-cell chromatin structure by Cryo-FIB and Cryo-ET.Curr Opin Struct Biol. 2025 Jun;92:103060. doi: 10.1016/j.sbi.2025.103060. Epub 2025 May 10. Curr Opin Struct Biol. 2025. PMID: 40349511 Free PMC article. Review.
-
High-throughput cryo-electron tomography enables multiscale visualization of the inner life of microbes.Curr Opin Struct Biol. 2025 Aug;93:103065. doi: 10.1016/j.sbi.2025.103065. Epub 2025 May 16. Curr Opin Struct Biol. 2025. PMID: 40381356 Free PMC article. Review.
References
-
- Bai XC, McMullan G & Scheres SH How cryo-EM is revolutionizing structural biology. Trends Biochem Sci 40, 49–57 (2015). - PubMed
-
- Murata K & Wolf M Cryo-electron microscopy for structural analysis of dynamic biological macromolecules. Biochim Biophys Acta Gen Subj 1862, 324–334 (2018). - PubMed
-
- Punjani A & Fleet DJ 3D variability analysis: Resolving continuous flexibility and discrete heterogeneity from single particle cryo-EM. J Struct Biol 213, 107702 (2021). - PubMed
MeSH terms
Grants and funding
- R01-GM144542/U.S. Department of Health & Human Services | NIH | National Institute of General Medical Sciences (NIGMS)
- 2046778/National Science Foundation (NSF)
- R01 GM144542/GM/NIGMS NIH HHS/United States
- 5T32-GM007287/U.S. Department of Health & Human Services | NIH | National Institute of General Medical Sciences (NIGMS)
- T32 GM007287/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
