

Hemispheric dominance in HVC is experience-dependent in juvenile male zebra finches
- PMID: 38461197
- PMCID: PMC10924951
- DOI: 10.1038/s41598-024-55987-6
Hemispheric dominance in HVC is experience-dependent in juvenile male zebra finches
Abstract
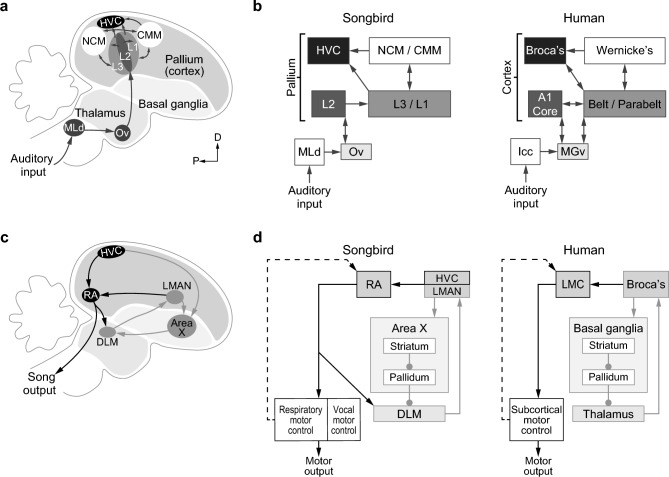
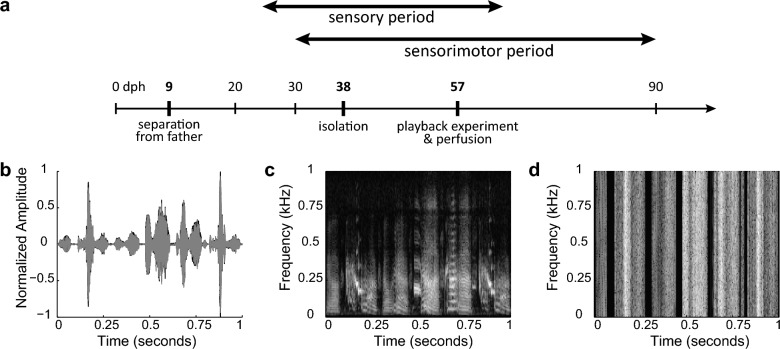
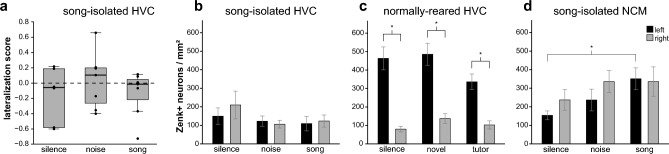
Juvenile male zebra finches (Taeniopygia guttata) must be exposed to an adult tutor during a sensitive period to develop normal adult song. The pre-motor nucleus HVC (acronym used as a proper name), plays a critical role in song learning and production (cf. Broca's area in humans). In the human brain, left-side hemispheric dominance in some language regions is positively correlated with proficiency in linguistic skills. However, it is unclear whether this pattern depends upon language learning, develops with normal maturation of the brain, or is the result of pre-existing functional asymmetries. In juvenile zebra finches, even though both left and right HVC contribute to song production, baseline molecular activity in HVC is left-dominant. To test if HVC exhibits hemispheric dominance prior to song learning, we raised juvenile males in isolation from adult song and measured neuronal activity in the left and right HVC upon first exposure to an auditory stimulus. Activity in the HVC was measured using the immediate early gene (IEG) zenk (acronym for zif-268, egr-1, NGFI-a, and krox-24) as a marker for neuronal activity. We found that neuronal activity in the HVC of juvenile male zebra finches is not lateralized when raised in the absence of adult song, while normally-reared juvenile birds are left-dominant. These findings show that there is no pre-existing asymmetry in the HVC prior to song exposure, suggesting that lateralization of the song system depends on learning through early exposure to adult song and subsequent song-imitation practice.
Keywords: Broca’s area; Language learning; Lateralization; Song learning; Zenk.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Experience-Dependent Intrinsic Plasticity During Auditory Learning.J Neurosci. 2019 Feb 13;39(7):1206-1221. doi: 10.1523/JNEUROSCI.1036-18.2018. Epub 2018 Dec 12. J Neurosci. 2019. PMID: 30541908 Free PMC article.
-
Human-like brain hemispheric dominance in birdsong learning.Proc Natl Acad Sci U S A. 2012 Jul 31;109(31):12782-7. doi: 10.1073/pnas.1207207109. Epub 2012 Jul 16. Proc Natl Acad Sci U S A. 2012. PMID: 22802637 Free PMC article.
-
Mirrored patterns of lateralized neuronal activation reflect old and new memories in the avian auditory cortex.Neuroscience. 2016 Aug 25;330:395-402. doi: 10.1016/j.neuroscience.2016.06.009. Epub 2016 Jun 8. Neuroscience. 2016. PMID: 27288718 Free PMC article.
-
The sensitive period for auditory-vocal learning in the zebra finch: Consequences of limited-model availability and multiple-tutor paradigms on song imitation.Behav Processes. 2019 Jun;163:5-12. doi: 10.1016/j.beproc.2017.07.007. Epub 2017 Jul 23. Behav Processes. 2019. PMID: 28743517 Free PMC article. Review.
-
Birdsong memory and the brain: in search of the template.Neurosci Biobehav Rev. 2015 Mar;50:41-55. doi: 10.1016/j.neubiorev.2014.11.019. Epub 2014 Nov 29. Neurosci Biobehav Rev. 2015. PMID: 25459663 Review.
Cited by
-
The songbird connectome (OSCINE-NET.ORG): structure-function organization beyond the canonical vocal control network.BMC Neurosci. 2024 Dec 27;25(1):79. doi: 10.1186/s12868-024-00919-3. BMC Neurosci. 2024. PMID: 39731002 Free PMC article.
-
Evaluating alpha-synuclein proteinopathy and consequences for birdsong in zebra finch basal ganglia area X.Behav Brain Res. 2025 Sep 13;493:115698. doi: 10.1016/j.bbr.2025.115698. Epub 2025 Jun 10. Behav Brain Res. 2025. PMID: 40505974
References
-
- Rogers LJ. Lateralization in vertebrates: Its early evolution, general pattern, and development. Adv. Study Behav. 2002;31:107–161. doi: 10.1016/S0065-3454(02)80007-9. - DOI
-
- Frasnelli E, Vallortigara G. Individual-level and population-level lateralization: Two sides of the same coin. Symmetry-Basel. 2018;10:12. doi: 10.3390/sym10120739. - DOI
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
