

This is a preprint.
Hachiman is a genome integrity sensor
- PMID: 38464307
- PMCID: PMC10925250
- DOI: 10.1101/2024.02.29.582594
Hachiman is a genome integrity sensor
Update in
-
Genome integrity sensing by the broad-spectrum Hachiman antiphage defense complex.Cell. 2024 Nov 27;187(24):6914-6928.e20. doi: 10.1016/j.cell.2024.09.020. Epub 2024 Oct 11. Cell. 2024. PMID: 39395413 Free PMC article.
Abstract
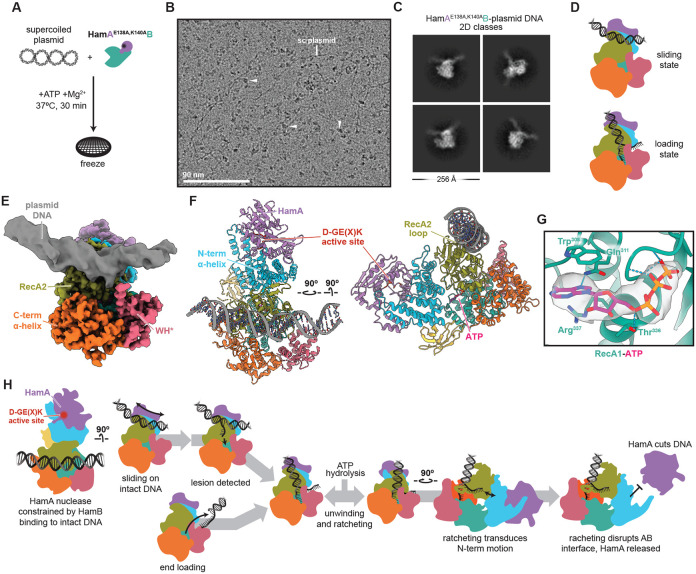
Hachiman is a broad-spectrum antiphage defense system of unknown function. We show here that Hachiman comprises a heterodimeric nuclease-helicase complex, HamAB. HamA, previously a protein of unknown function, is the effector nuclease. HamB is the sensor helicase. HamB constrains HamA activity during surveillance of intact dsDNA. When the HamAB complex detects DNA damage, HamB helicase activity liberates HamA, unleashing nuclease activity. Hachiman activation degrades all DNA in the cell, creating 'phantom' cells devoid of both phage and host DNA. We demonstrate Hachiman activation in the absence of phage by treatment with DNA-damaging agents, suggesting that Hachiman responds to aberrant DNA states. Phylogenetic similarities between the Hachiman helicase and eukaryotic enzymes suggest this bacterial immune system has been repurposed for diverse functions across all domains of life.
Conflict of interest statement
Competing interest statement J.A.D. is a co-founder of Caribou Biosciences, Editas Medicine, Intellia Therapeutics, Mammoth Biosciences and Scribe Therapeutics, and a director of Altos, Johnson & Johnson and Tempus. J.A.D. is a scientific advisor to Caribou Biosciences, Intellia Therapeutics, Mammoth Biosciences, Inari, Scribe Therapeutics, Felix Biosciences and Algen. J.A.D. also serves as Chief Science Advisor to Sixth Street and a Scientific Advisory Board member at The Column Group. J.A.D. conducts academic research projects sponsored by Roche and Apple Tree Partners. J.P. has an equity interest in Linnaeus Bioscience Incorporated and receives income. The terms of this arrangement have been reviewed and approved by the University of California, San Diego, in accordance with its conflict-of-interest policies.
Figures






References
-
- Singleton M. R., Dillingham M. S. & Wigley D. B. Structure and 1. Singleton M. R., Dillingham M. S. & Wigley D. B. Structure and mechanism of helicases and nucleic acid translocases. Annu. Rev. Biochem. 76, 23–50 (2007). - PubMed
-
- Pyle A. M. Translocation and unwinding mechanisms of RNA and DNA helicases. Annu. Rev. Biophys. 37, 317–336 (2008). - PubMed
-
- Bleichert F., Botchan M. R. & Berger J. M. Mechanisms for initiating cellular DNA replication. Science 355, (2017). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources