

Structure of RADX and mechanism for regulation of RAD51 nucleofilaments
- PMID: 38466836
- PMCID: PMC10962997
- DOI: 10.1073/pnas.2316491121
Structure of RADX and mechanism for regulation of RAD51 nucleofilaments
Abstract
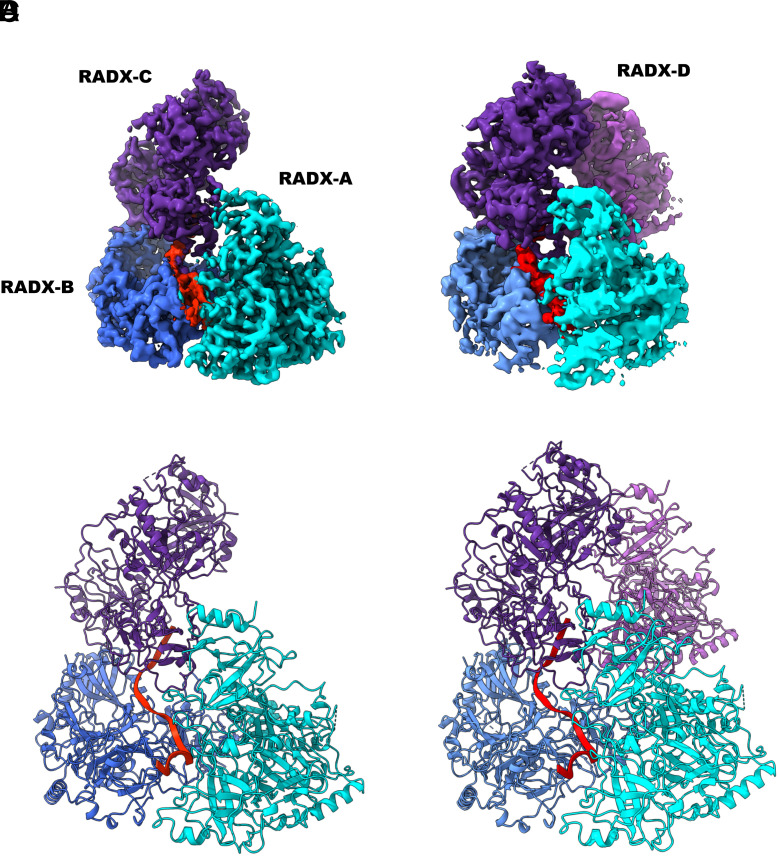
Replication fork reversal is a fundamental process required for resolution of encounters with DNA damage. A key step in the stabilization and eventual resolution of reversed forks is formation of RAD51 nucleoprotein filaments on exposed single strand DNA (ssDNA). To avoid genome instability, RAD51 filaments are tightly controlled by a variety of positive and negative regulators. RADX (RPA-related RAD51-antagonist on the X chromosome) is a recently discovered negative regulator that binds tightly to ssDNA, directly interacts with RAD51, and regulates replication fork reversal and stabilization in a context-dependent manner. Here, we present a structure-based investigation of RADX's mechanism of action. Mass photometry experiments showed that RADX forms multiple oligomeric states in a concentration-dependent manner, with a predominance of trimers in the presence of ssDNA. The structure of RADX, which has no structurally characterized orthologs, was determined ab initio by cryo-electron microscopy (cryo-EM) from maps in the 2 to 4 Å range. The structure reveals the molecular basis for RADX oligomerization and the coupled multi-valent binding of ssDNA binding. The interaction of RADX with RAD51 filaments was imaged by negative stain EM, which showed a RADX oligomer at the end of filaments. Based on these results, we propose a model in which RADX functions by capping and restricting the end of RAD51 filaments.
Keywords: DNA replication; RAD51; cryo-EM; replication fork.
Conflict of interest statement
Competing interests statement:The authors declare no competing interest.
Figures






Update of
-
Structure of RADX and mechanism for regulation of RAD51 nucleofilaments.bioRxiv [Preprint]. 2023 Sep 20:2023.09.19.558089. doi: 10.1101/2023.09.19.558089. bioRxiv. 2023. Update in: Proc Natl Acad Sci U S A. 2024 Mar 19;121(12):e2316491121. doi: 10.1073/pnas.2316491121. PMID: 37786681 Free PMC article. Updated. Preprint.
References
-
- Berti M., Cortez D., Lopes M., The plasticity of DNA replication forks in response to clinically relevant genotoxic stress. Nat. Rev. Mol. Cell Biol. 21, 633–651 (2020). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
