

Vascular ATGL-dependent lipolysis and the activation of cPLA2-PGI2 pathway protect against postprandial endothelial dysfunction
- PMID: 38467757
- PMCID: PMC10927860
- DOI: 10.1007/s00018-024-05167-6
Vascular ATGL-dependent lipolysis and the activation of cPLA2-PGI2 pathway protect against postprandial endothelial dysfunction
Abstract
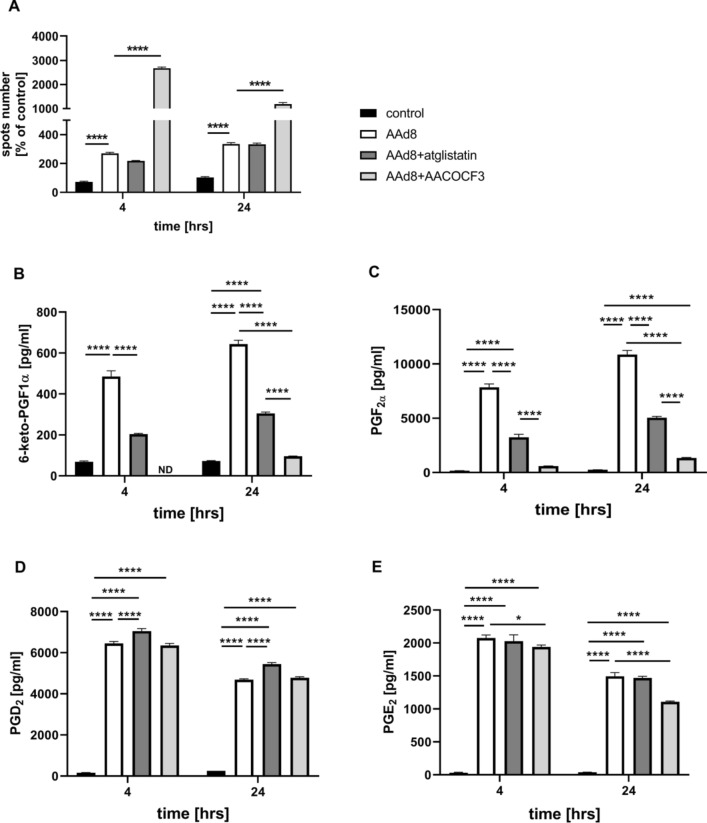
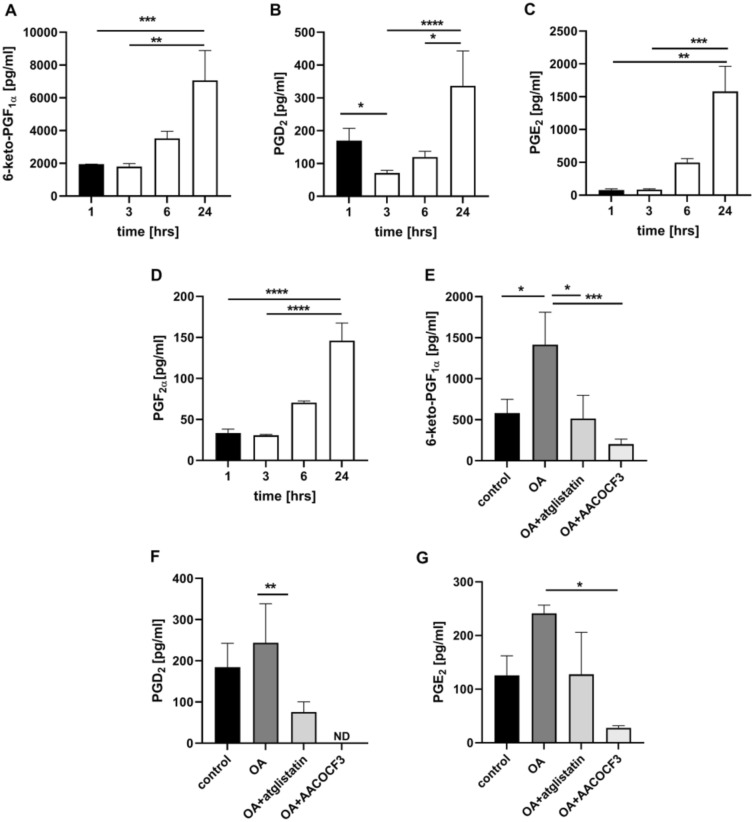
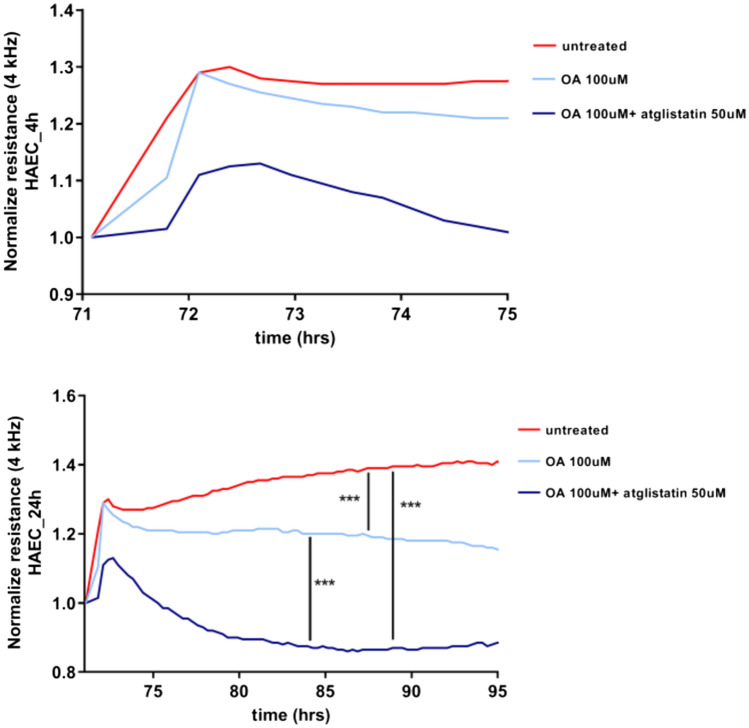
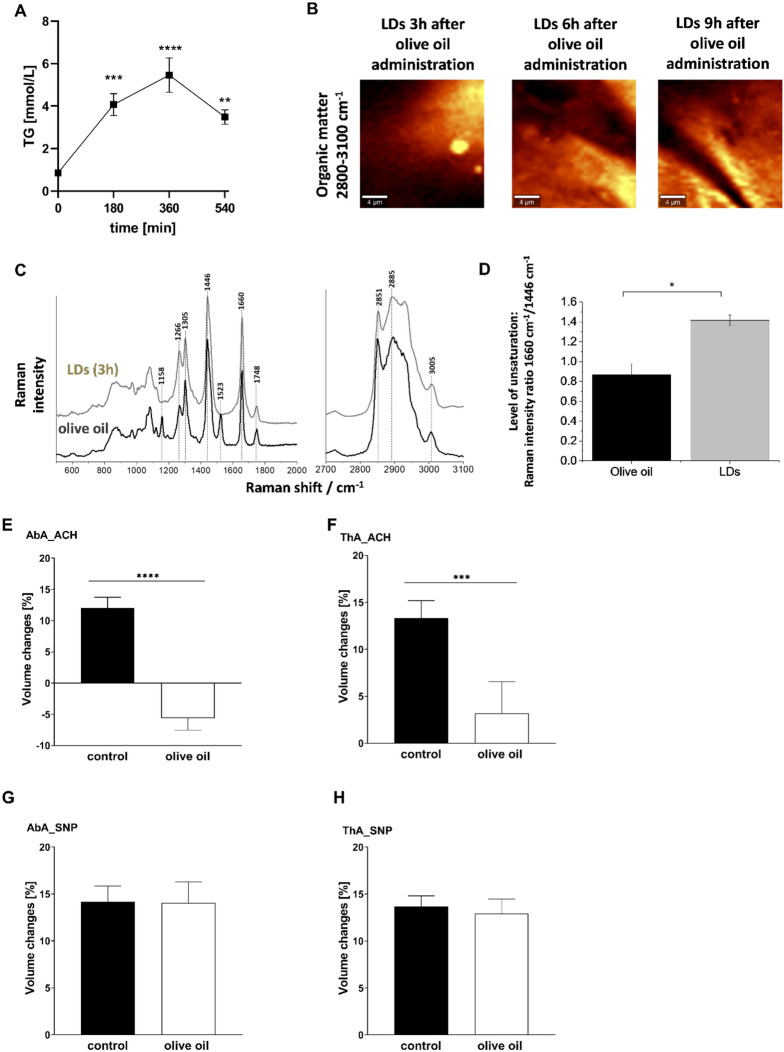
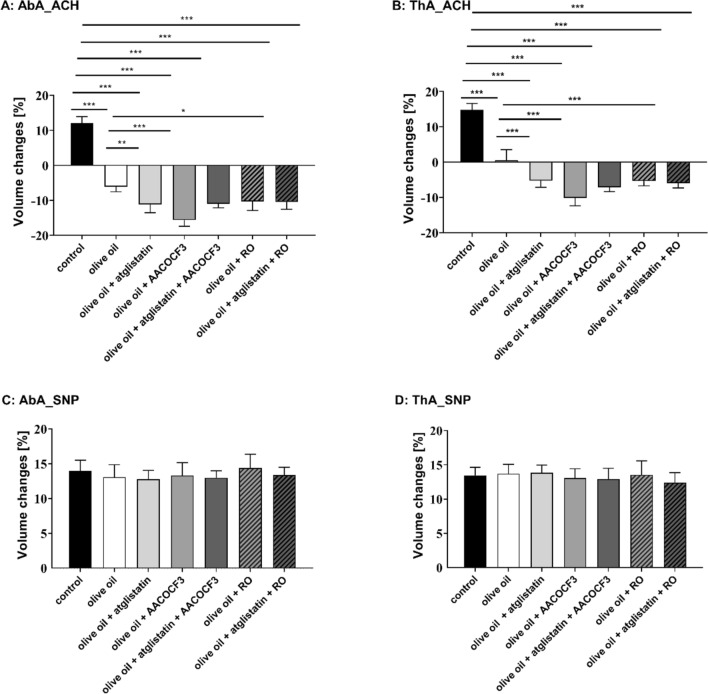
Adipose triglyceride lipase (ATGL) is involved in lipolysis and displays a detrimental pathophysiological role in cardio-metabolic diseases. However, the organo-protective effects of ATGL-induced lipolysis were also suggested. The aim of this work was to characterize the function of lipid droplets (LDs) and ATGL-induced lipolysis in the regulation of endothelial function. ATGL-dependent LDs hydrolysis and cytosolic phospholipase A2 (cPLA2)-derived eicosanoids production were studied in the aorta, endothelial and smooth muscle cells exposed to exogenous oleic acid (OA) or arachidonic acid (AA). Functional effects of ATGL-dependent lipolysis and subsequent activation of cPLA2/PGI2 pathway were also studied in vivo in relation to postprandial endothelial dysfunction.The formation of LDs was invariably associated with elevated production of endogenous AA-derived prostacyclin (PGI2). In the presence of the inhibitor of ATGL or the inhibitor of cytosolic phospholipase A2, the production of eicosanoids was reduced, with a concomitant increase in the number of LDs. OA administration impaired endothelial barrier integrity in vitro that was further impaired if OA was given together with ATGL inhibitor. Importantly, in vivo, olive oil induced postprandial endothelial dysfunction that was significantly deteriorated by ATGL inhibition, cPLA2 inhibition or by prostacyclin (IP) receptor blockade.In summary, vascular LDs formation induced by exogenous AA or OA was associated with ATGL- and cPLA2-dependent PGI2 production from endogenous AA. The inhibition of ATGL resulted in an impairment of endothelial barrier function in vitro. The inhibition of ATGL-cPLA2-PGI2 dependent pathway resulted in the deterioration of endothelial function upon exposure to olive oil in vivo. In conclusion, vascular ATGL-cPLA2-PGI2 dependent pathway activated by lipid overload and linked to LDs formation in endothelium and smooth muscle cells has a vasoprotective role by counterbalancing detrimental effects of lipid overload on endothelial function.
Keywords: ATGL; Atglistatin; Endothelial-induced vasodilation; Endothelium; Lipid droplets; Lipolysis.
© 2024. The Author(s).
Conflict of interest statement
Not applicable.
Figures



Similar articles
-
Vascular lipid droplets formed in response to TNF, hypoxia, or OA: biochemical composition and prostacyclin generation.J Lipid Res. 2023 May;64(5):100355. doi: 10.1016/j.jlr.2023.100355. Epub 2023 Mar 17. J Lipid Res. 2023. PMID: 36934842 Free PMC article.
-
Adipose triglyceride lipase regulates eicosanoid production in activated human mast cells.J Lipid Res. 2014 Dec;55(12):2471-8. doi: 10.1194/jlr.M048553. Epub 2014 Aug 11. J Lipid Res. 2014. PMID: 25114172 Free PMC article.
-
Reduced expression of adipose triglyceride lipase decreases arachidonic acid release and prostacyclin secretion in human aortic endothelial cells.Arch Physiol Biochem. 2017 Oct;123(4):249-253. doi: 10.1080/13813455.2017.1309052. Epub 2017 Apr 3. Arch Physiol Biochem. 2017. PMID: 28368219 Free PMC article.
-
[Adipose triglyceride lipase regulates adipocyte lipolysis].Sheng Li Ke Xue Jin Zhan. 2008 Jan;39(1):10-4. Sheng Li Ke Xue Jin Zhan. 2008. PMID: 18357681 Review. Chinese.
-
Mast cells: from lipid droplets to lipid mediators.Clin Sci (Lond). 2013 Aug;125(3):121-30. doi: 10.1042/CS20120602. Clin Sci (Lond). 2013. PMID: 23577635 Free PMC article. Review.
Cited by
-
Endothelial deletion of adipose triglyceride lipase protects against heart failure with preserved ejection fraction.JCI Insight. 2025 Mar 10;10(5):e187145. doi: 10.1172/jci.insight.187145. JCI Insight. 2025. PMID: 40059832 Free PMC article. No abstract available.
References
-
- Schweiger M, Romauch M, Schreiber R, Grabner GF, Hütter S, Kotzbeck P, Benedikt P, Eichmann TO, Yamada S, Knittelfelder O, Diwoky C, Doler C, Mayer N, De Cecco W, Breinbauer R, Zimmermann R, Zechner R. Pharmacological inhibition of adipose triglyceride lipase corrects high-fat diet-induced insulin resistance and hepatosteatosis in mice. Nat Commun. 2017;8:14859. doi: 10.1038/ncomms14859. - DOI - PMC - PubMed
-
- Thiele A, Luettges K, Ritter D, Beyhoff N, Smeir E, Grune J, Steinhoff JS, Schupp M, Klopfleisch R, Rothe M, Wilck N, Bartolomaeus H, Migglautsch AK, Breinbauer R, Kershaw EE, Grabner GF, Zechner R, Kintscher U, Foryst-Ludwig A. Pharmacological inhibition of adipose tissue adipose triglyceride lipase by Atglistatin prevents catecholamine-induced myocardial damage. Cardiovasc Res. 2021;118:2488. doi: 10.1093/cvr/cvab182. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
