

Rapid alkalinization factor 22 has a structural and signalling role in root hair cell wall assembly
- PMID: 38467800
- PMCID: PMC11494403
- DOI: 10.1038/s41477-024-01637-8
Rapid alkalinization factor 22 has a structural and signalling role in root hair cell wall assembly
Erratum in
-
Author Correction: Rapid alkalinization factor 22 has a structural and signalling role in root hair cell wall assembly.Nat Plants. 2024 Aug;10(8):1267. doi: 10.1038/s41477-024-01765-1. Nat Plants. 2024. PMID: 39043822 No abstract available.
Abstract
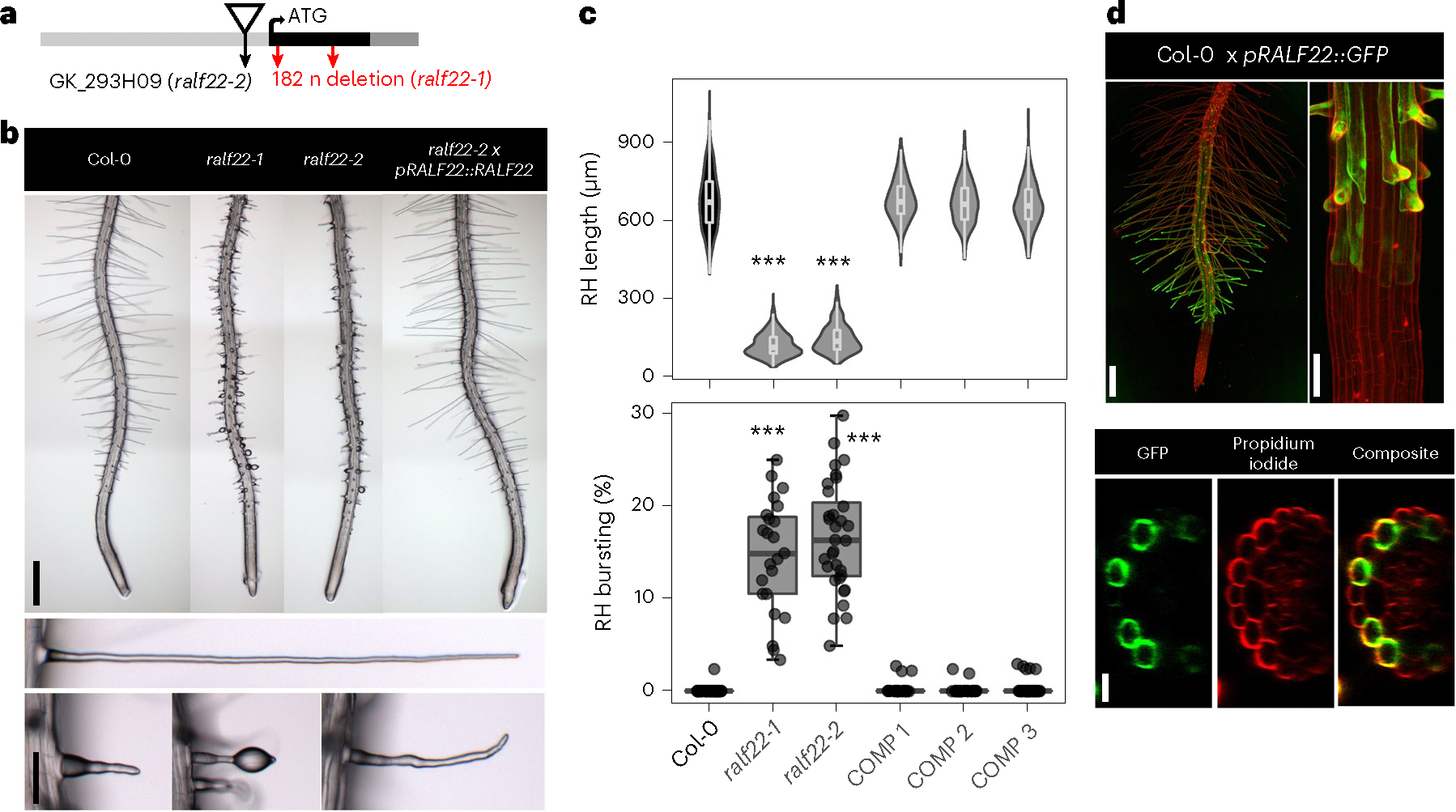
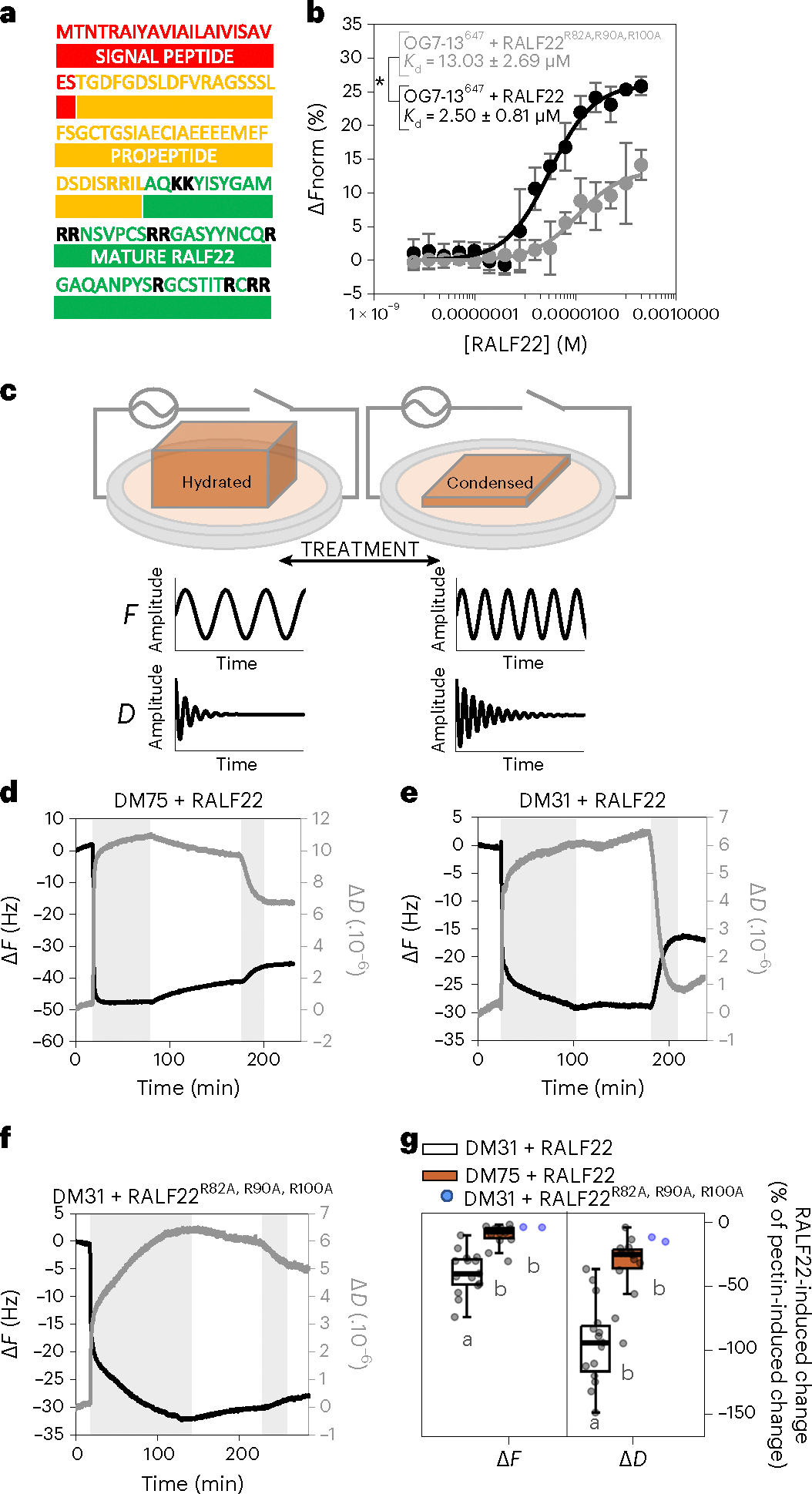
Pressurized cells with strong walls make up the hydrostatic skeleton of plants. Assembly and expansion of such stressed walls depend on a family of secreted RAPID ALKALINIZATION FACTOR (RALF) peptides, which bind both a membrane receptor complex and wall-localized LEUCINE-RICH REPEAT EXTENSIN (LRXs) in a mutually exclusive way. Here we show that, in root hairs, the RALF22 peptide has a dual structural and signalling role in cell expansion. Together with LRX1, it directs the compaction of charged pectin polymers at the root hair tip into periodic circumferential rings. Free RALF22 induces the formation of a complex with LORELEI-LIKE-GPI-ANCHORED PROTEIN 1 and FERONIA, triggering adaptive cellular responses. These findings show how a peptide simultaneously functions as a structural component organizing cell wall architecture and as a feedback signalling molecule that regulates this process depending on its interaction partners. This mechanism may also underlie wall assembly and expansion in other plant cell types.
© 2024. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures





References
-
- Kjellén L & Lindahl U Specificity of glycosaminoglycan–protein interactions. Curr. Opin. Struct. Biol. 50, 101–108 (2018). - PubMed
-
- Coen E & Cosgrove DJ The mechanics of plant morphogenesis. Science 379, eade8055 (2023). - PubMed
-
- Levesque-Tremblay G, Pelloux J, Braybrook SA & Müller K Tuning of pectin methylesterification: consequences for cell wall biomechanics and development. Planta 242, 791–811 (2015). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 GM131043/GM/NIGMS NIH HHS/United States
- AUHA-15-12/Fonds Wetenschappelijk Onderzoek (Research Foundation Flanders)
- I003420N/Fonds Wetenschappelijk Onderzoek (Research Foundation Flanders)
- 1225120N/Fonds Wetenschappelijk Onderzoek (Research Foundation Flanders)
- AUHA-09-001/Fonds Wetenschappelijk Onderzoek (Research Foundation Flanders)
LinkOut - more resources
Full Text Sources
Research Materials
