

P. aeruginosa CtpA protease adopts a novel activation mechanism to initiate the proteolytic process
- PMID: 38467832
- PMCID: PMC11021448
- DOI: 10.1038/s44318-024-00069-6
P. aeruginosa CtpA protease adopts a novel activation mechanism to initiate the proteolytic process
Abstract
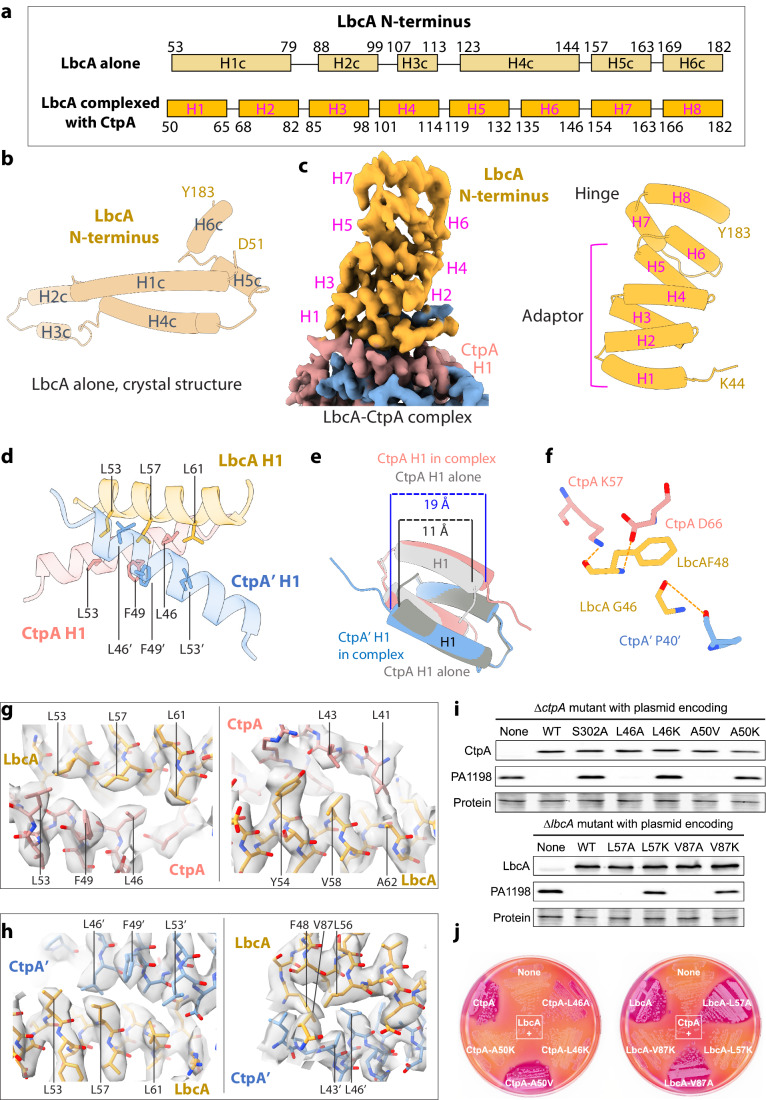
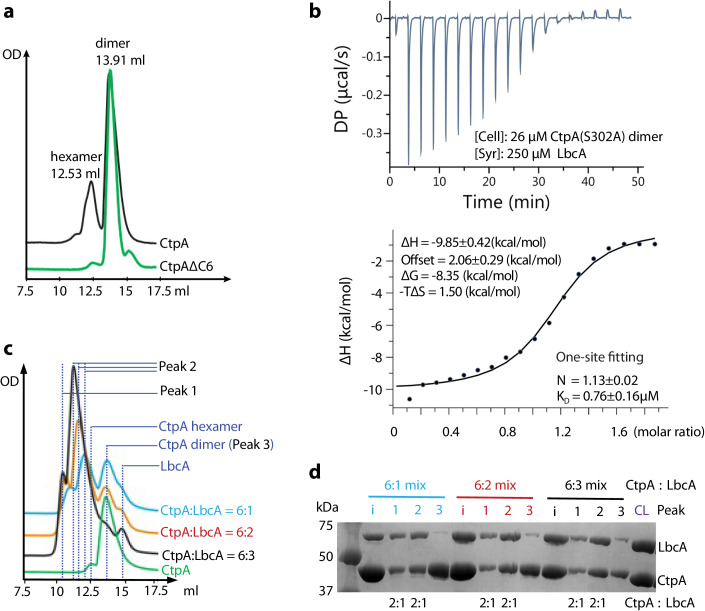
During bacterial cell growth, hydrolases cleave peptide cross-links between strands of the peptidoglycan sacculus to allow new strand insertion. The Pseudomonas aeruginosa carboxyl-terminal processing protease (CTP) CtpA regulates some of these hydrolases by degrading them. CtpA assembles as an inactive hexamer composed of a trimer-of-dimers, but its lipoprotein binding partner LbcA activates CtpA by an unknown mechanism. Here, we report the cryo-EM structures of the CtpA-LbcA complex. LbcA has an N-terminal adaptor domain that binds to CtpA, and a C-terminal superhelical tetratricopeptide repeat domain. One LbcA molecule attaches to each of the three vertices of a CtpA hexamer. LbcA triggers relocation of the CtpA PDZ domain, remodeling of the substrate binding pocket, and realignment of the catalytic residues. Surprisingly, only one CtpA molecule in a CtpA dimer is activated upon LbcA binding. Also, a long loop from one CtpA dimer inserts into a neighboring dimer to facilitate the proteolytic activity. This work has revealed an activation mechanism for a bacterial CTP that is strikingly different from other CTPs that have been characterized structurally.
Keywords: Pseudomonas aeruginosa; Cell Wall; Peptidoglycan Hydrolases; Protease Activation; Protein Complex.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures






Similar articles
-
Pseudomonas aeruginosa C-Terminal Processing Protease CtpA Assembles into a Hexameric Structure That Requires Activation by a Spiral-Shaped Lipoprotein-Binding Partner.mBio. 2022 Feb 22;13(1):e0368021. doi: 10.1128/mbio.03680-21. Epub 2022 Jan 18. mBio. 2022. PMID: 35038915 Free PMC article.
-
A Proteolytic Complex Targets Multiple Cell Wall Hydrolases in Pseudomonas aeruginosa.mBio. 2018 Jul 17;9(4):e00972-18. doi: 10.1128/mBio.00972-18. mBio. 2018. PMID: 30018106 Free PMC article.
-
Direct and Indirect Interactions Promote Complexes of the Lipoprotein LbcA, the CtpA Protease and Its Substrates, and Other Cell Wall Proteins in Pseudomonas aeruginosa.J Bacteriol. 2021 Nov 19;203(24):e0039321. doi: 10.1128/JB.00393-21. Epub 2021 Sep 27. J Bacteriol. 2021. PMID: 34570626 Free PMC article.
-
Bacterial Carboxyl-Terminal Processing Proteases Play Critical Roles in the Cell Envelope and Beyond.J Bacteriol. 2022 Apr 19;204(4):e0062821. doi: 10.1128/jb.00628-21. Epub 2022 Mar 16. J Bacteriol. 2022. PMID: 35293777 Free PMC article. Review.
-
[Mechanism of action of proteinases of Pseudomonas aeruginosa].Pathol Biol (Paris). 1990 Dec;38(10):968-74. Pathol Biol (Paris). 1990. PMID: 2127091 Review. French.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
