

Control of three-carbon amino acid homeostasis by promiscuous importers and exporters in Bacillus subtilis: role of the "sleeping beauty" amino acid exporters
- PMID: 38470260
- PMCID: PMC11005379
- DOI: 10.1128/mbio.03456-23
Control of three-carbon amino acid homeostasis by promiscuous importers and exporters in Bacillus subtilis: role of the "sleeping beauty" amino acid exporters
Abstract
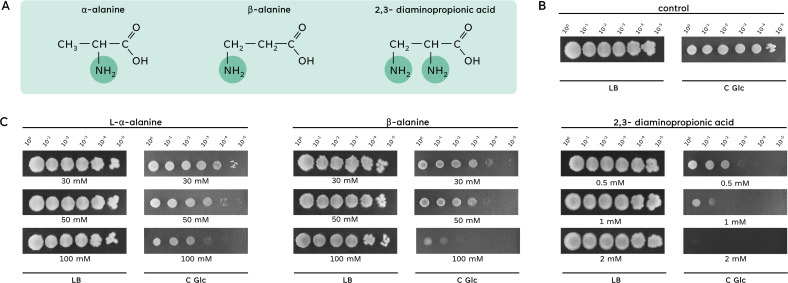
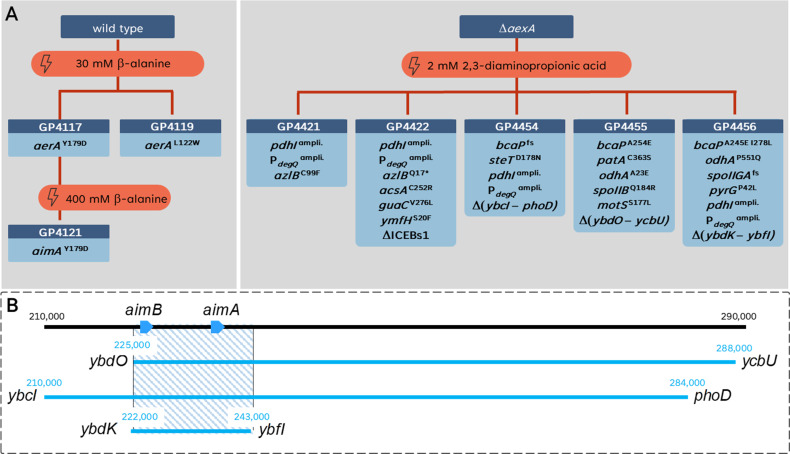
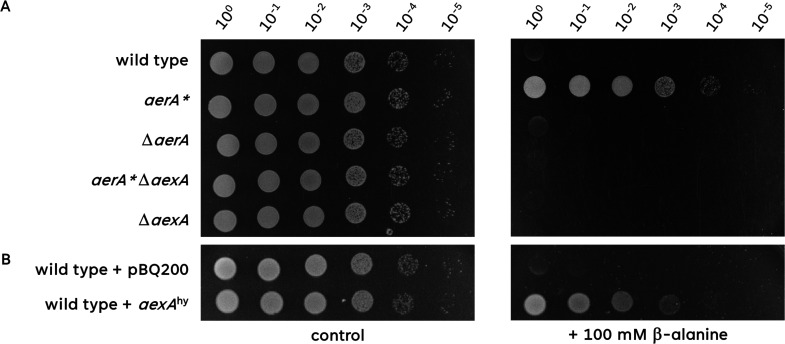
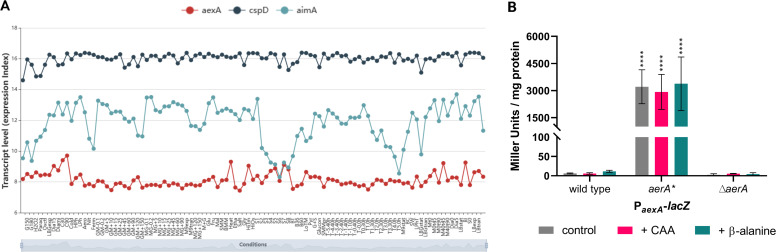
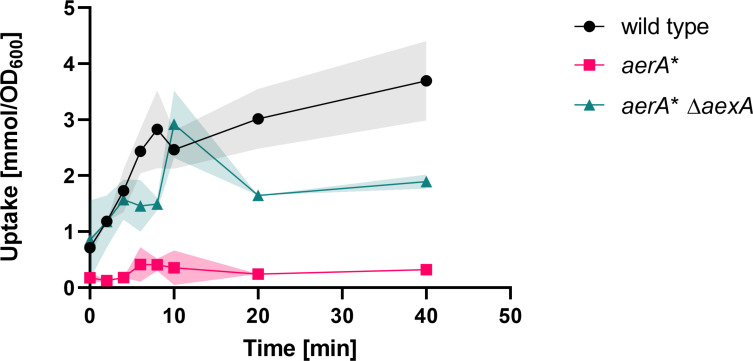
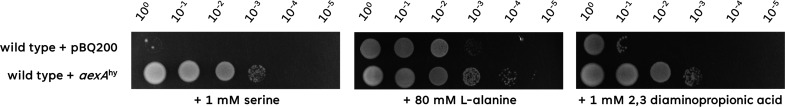
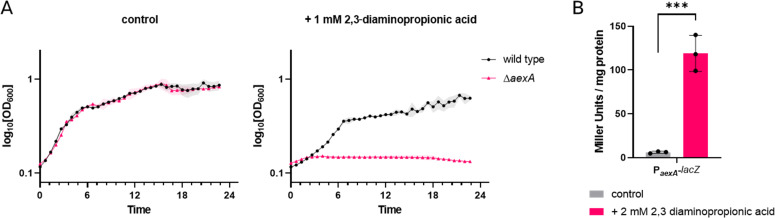
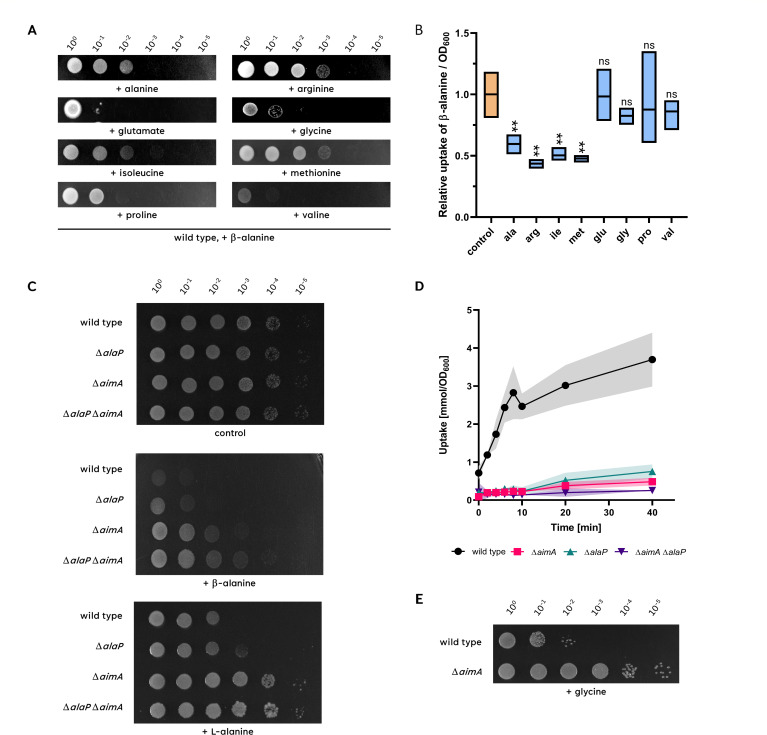
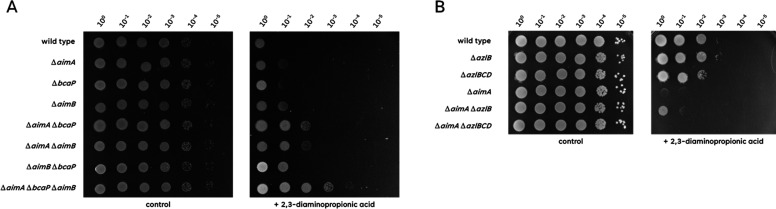
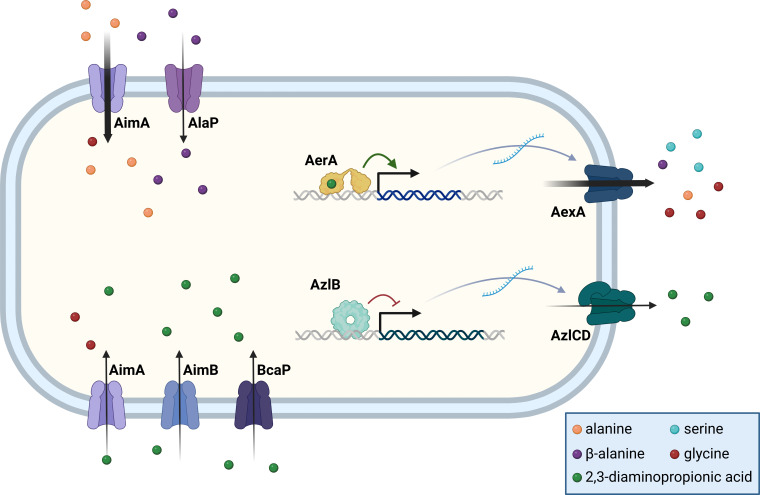
The Gram-positive model bacterium Bacillus subtilis can acquire amino acids by import, de novo biosynthesis, or degradation of proteins and peptides. The accumulation of several amino acids inhibits the growth of B. subtilis, probably due to misincorporation into cellular macromolecules such as proteins or peptidoglycan or due to interference with other amino acid biosynthetic pathways. Here, we studied the adaptation of B. subtilis to toxic concentrations of the three-carbon amino acids L-alanine, β-alanine, and 2,3-diaminopropionic acid, as well as the two-carbon amino acid glycine. Resistance to the non-proteinogenic amino acid β-alanine, which is a precursor for coenzyme A biosynthesis, is achieved by mutations that either activate a cryptic amino acid exporter, AexA (previously YdeD), or inactivate the amino acid importers AimA, AimB (previously YbxG), and BcaP. The aexA gene is very poorly expressed under most conditions studied. However, mutations affecting the transcription factor AerA (previously YdeC) can result in strong constitutive aexA expression. AexA is the first characterized member of a group of amino acid exporters in B. subtilis, which are all very poorly expressed. Therefore, we suggest to call this group "sleeping beauty amino acid exporters." 2,3-Diaminopropionic acid can also be exported by AexA, and this amino acid also seems to be a natural substrate of AerA/AexA, as it can cause a slight but significant induction of aexA expression, and AexA also provides some natural resistance toward 2,3-diaminopropionic acid. Moreover, our work shows how low-specificity amino acid transporters contribute to amino acid homeostasis in B. subtilis.IMPORTANCEEven though Bacillus subtilis is one of the most-studied bacteria, amino acid homeostasis in this organism is not fully understood. We have identified import and export systems for the C2 and C3 amino acids. Our work demonstrates that the responsible amino acid permeases contribute in a rather promiscuitive way to amino acid uptake. In addition, we have discovered AexA, the first member of a group of very poorly expressed amino acid exporters in B. subtilis that we call "sleeping beauty amino acid exporters." The expression of these transporters is typically triggered by mutations in corresponding regulator genes that are acquired upon exposure to toxic amino acids. These exporters are ubiquitous in all domains of life. It is tempting to speculate that many of them are not expressed until the cells experience selective pressure by toxic compounds, and they protect the cells from rare but potentially dangerous encounters with such compounds.
Keywords: Bacillus subtilis; amino acid export; amino acid uptake; diaminopropionic acid; sleeping beauty amino acid exporters; β-alanine.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Control of asparagine homeostasis in Bacillus subtilis: identification of promiscuous amino acid importers and exporters.J Bacteriol. 2024 Feb 22;206(2):e0042023. doi: 10.1128/jb.00420-23. Epub 2024 Jan 9. J Bacteriol. 2024. PMID: 38193659 Free PMC article.
-
How To Deal with Toxic Amino Acids: the Bipartite AzlCD Complex Exports Histidine in Bacillus subtilis.J Bacteriol. 2022 Dec 20;204(12):e0035322. doi: 10.1128/jb.00353-22. Epub 2022 Nov 15. J Bacteriol. 2022. PMID: 36377869 Free PMC article.
-
Adaptation of Bacillus subtilis to Life at Extreme Potassium Limitation.mBio. 2017 Jul 5;8(4):e00861-17. doi: 10.1128/mBio.00861-17. mBio. 2017. PMID: 28679749 Free PMC article.
-
The ATP binding cassette (ABC) transport systems of Mycobacterium tuberculosis.FEMS Microbiol Rev. 2000 Oct;24(4):449-67. doi: 10.1111/j.1574-6976.2000.tb00550.x. FEMS Microbiol Rev. 2000. PMID: 10978546 Review.
-
The many roles of cyclic di-AMP to control the physiology of Bacillus subtilis.Microlife. 2023 Oct 20;4:uqad043. doi: 10.1093/femsml/uqad043. eCollection 2023. Microlife. 2023. PMID: 37954098 Free PMC article. Review.
Cited by
-
Coenzyme A biosynthesis in Bacillus subtilis: discovery of a novel precursor metabolite for salvage and its uptake system.mBio. 2024 Oct 16;15(10):e0177224. doi: 10.1128/mbio.01772-24. Epub 2024 Aug 28. mBio. 2024. PMID: 39194188 Free PMC article.
References
-
- Calogero S, Gardan R, Glaser P, Schweizer J, Rapoport G, Debarbouille M. 1994. RocR, a novel regulatory protein controlling arginine utilization in Bacillus subtilis, belongs to the NtrC/NifA family of transcriptional activators. J Bacteriol 176:1234–1241. doi:10.1128/jb.176.5.1234-1241.1994 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
