

Human Host Defense Peptide LL-37 Suppresses TNFα-Mediated Matrix Metalloproteinases MMP9 and MMP13 in Human Bronchial Epithelial Cells
- PMID: 38471488
- PMCID: PMC10997319
- DOI: 10.1159/000537775
Human Host Defense Peptide LL-37 Suppresses TNFα-Mediated Matrix Metalloproteinases MMP9 and MMP13 in Human Bronchial Epithelial Cells
Abstract
Introduction: TNFα-inducible matrix metalloproteinases play a critical role in the process of airway remodeling in respiratory inflammatory disease including asthma. The cationic host defense peptide LL-37 is elevated in the lungs during airway inflammation. However, the impact of LL-37 on TNFα-driven processes is not well understood. Here, we examined the effect of LL-37 on TNFα-mediated responses in human bronchial epithelial cells (HBECs).
Methods: We used a slow off-rate modified aptamer-based proteomics approach to define the HBEC proteome altered in response to TNFα. Abundance of selected protein candidates and signaling intermediates was examined using immunoassays, ELISA and Western blots, and mRNA abundance was examined by qRT-PCR.
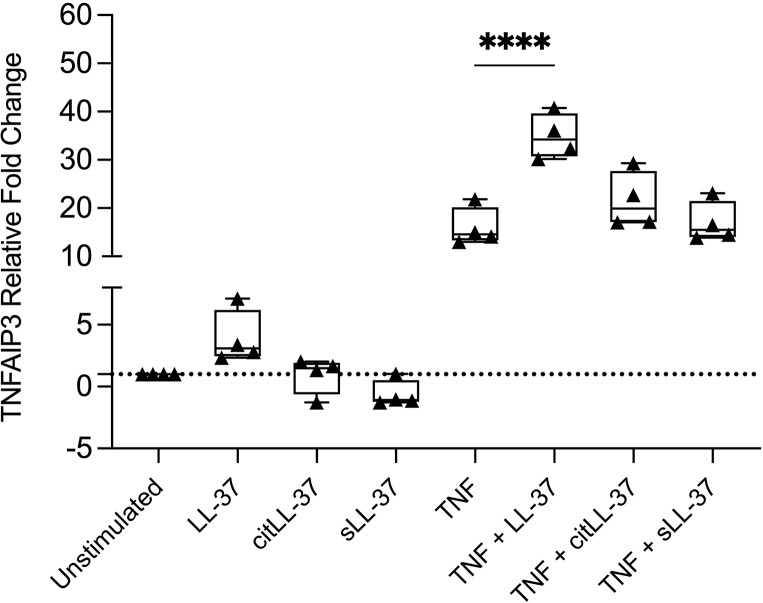
Results: Proteomics analysis revealed that 124 proteins were significantly altered, 12 proteins were enhanced by ≥2-fold compared to unstimulated cells, in response to TNFα. MMP9 was the topmost increased protein in response to TNFα, enhanced by ∼10-fold, and MMP13 was increased by ∼3-fold, compared to unstimulated cells. Furthermore, we demonstrated that LL-37 significantly suppressed TNFα-mediated MMP9 and MMP13 in HBEC. Mechanistic data revealed that TNFα-mediated MMP9 and MMP13 production is controlled by SRC kinase and that LL-37 enhances related upstream negative regulators, namely, phospho-AKT (T308) and TNFα-mediated TNFAIP3 or A20.
Conclusions: The findings of this study suggest that LL-37 may play a role in intervening in the process of airway remodeling in chronic inflammatory respiratory disease such as asthma.
Keywords: Airway remodeling; Cathelicidin; Host defense peptides; Inflammation; LL-37; Matrix metalloproteinases; TNFα.
© 2024 The Author(s). Published by S. Karger AG, Basel.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article.
Figures





References
-
- Banno A, Reddy AT, Lakshmi SP, Reddy RC. Bidirectional interaction of airway epithelial remodeling and inflammation in asthma. Clin Sci. 2020;134(9):1063–79. - PubMed
-
- Berry M, Brightling C, Pavord I, Wardlaw A. TNF-alpha in asthma. Curr Opin Pharmacol. 2007;7(3):279–82. - PubMed
-
- Berry MA, Hargadon B, Shelley M, Parker D, Shaw DE, Green RH, et al. . Evidence of a role of tumor necrosis factor alpha in refractory asthma. N Engl J Med. 2006;354(7):697–708. - PubMed
-
- Atkinson JJ, Senior RM. Matrix metalloproteinase-9 in lung remodeling. Am J Respir Cell Mol Biol. 2003;28(1):12–24. - PubMed
-
- Gras D, Chanez P, Vachier I, Petit A, Bourdin A. Bronchial epithelium as a target for innovative treatments in asthma. Pharmacol Ther. 2013;140(3):290–305. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
