

The underappreciated diversity of bile acid modifications
- PMID: 38471500
- PMCID: PMC12248420
- DOI: 10.1016/j.cell.2024.02.019
The underappreciated diversity of bile acid modifications
Abstract
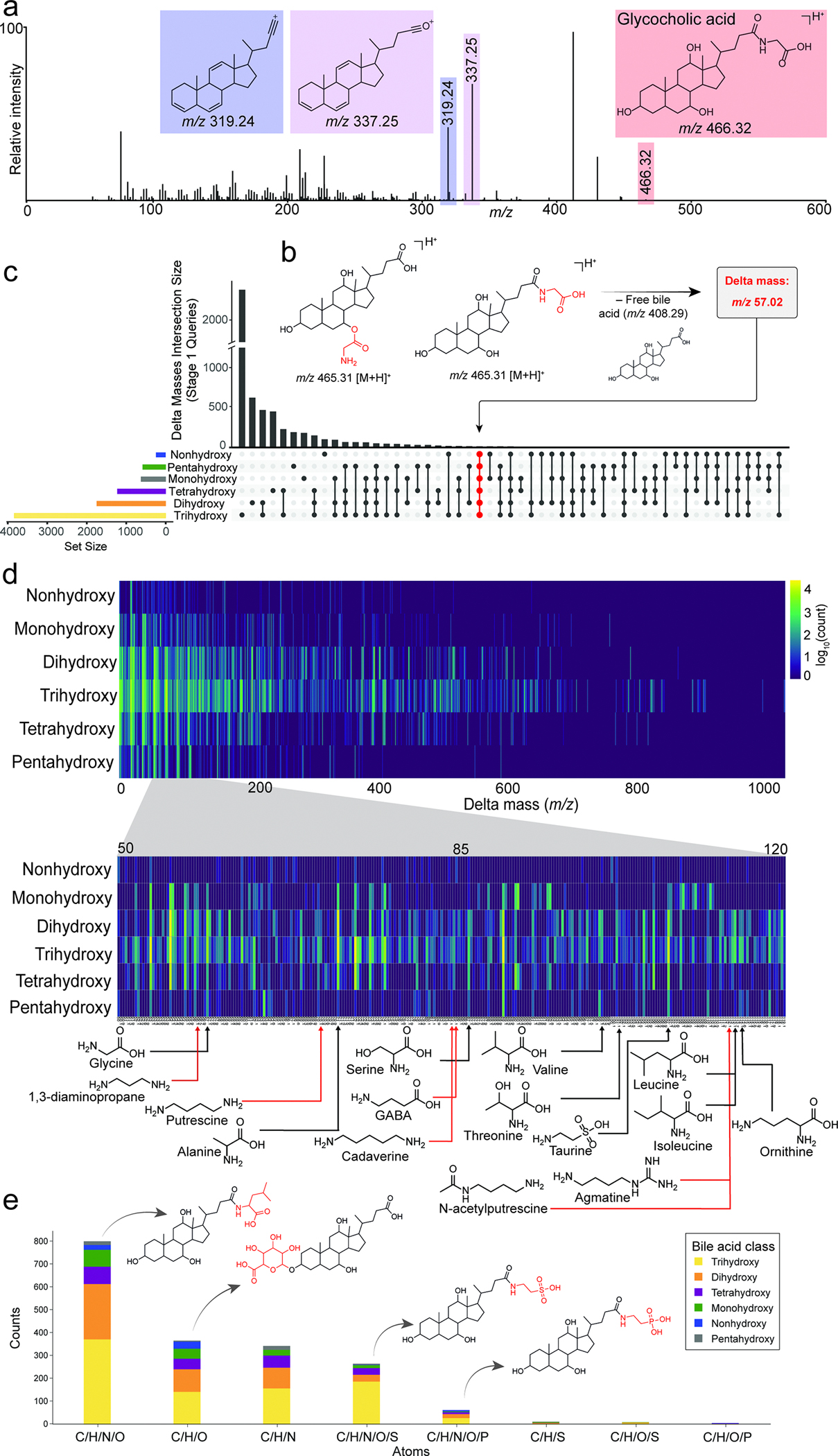
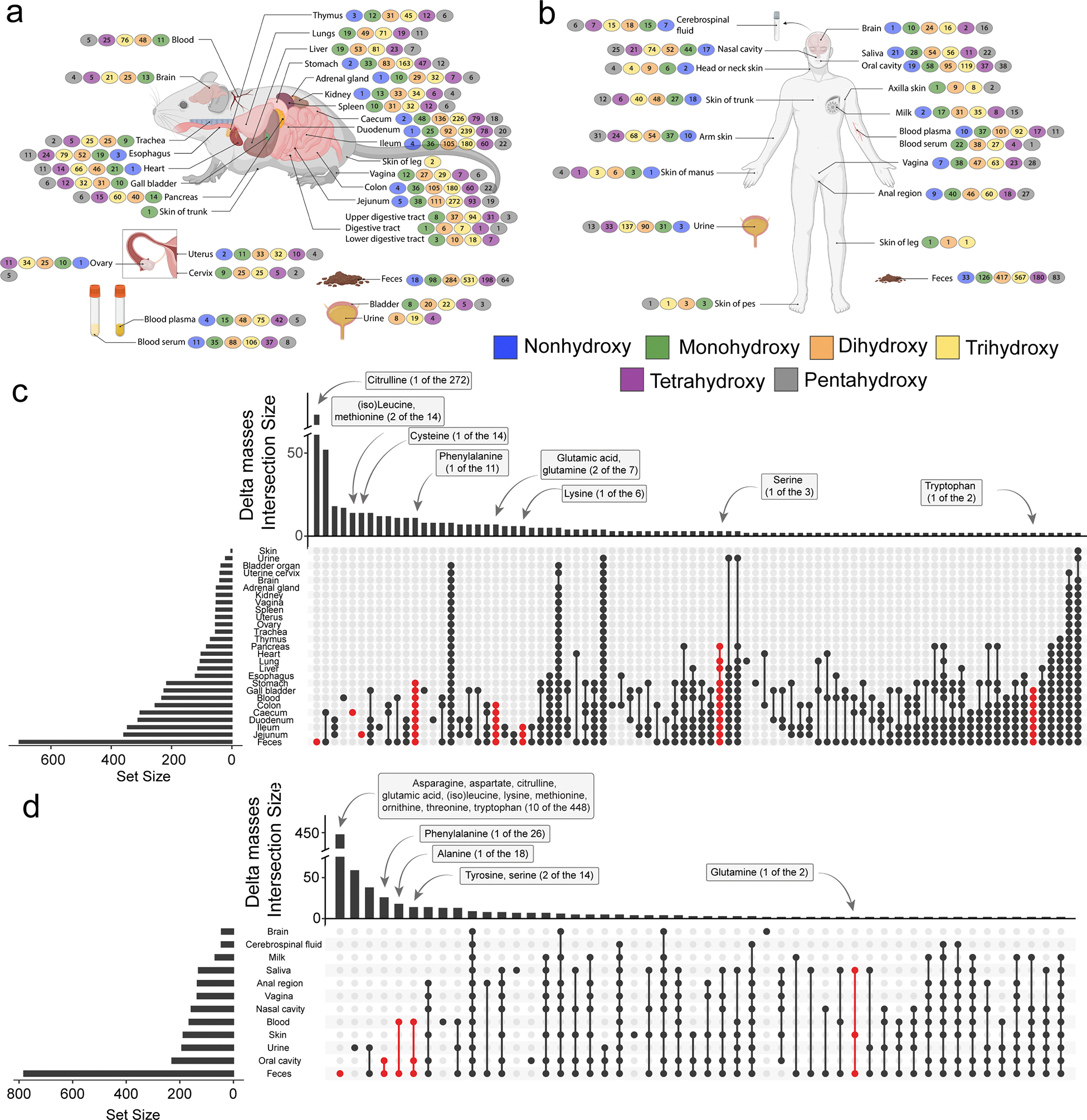
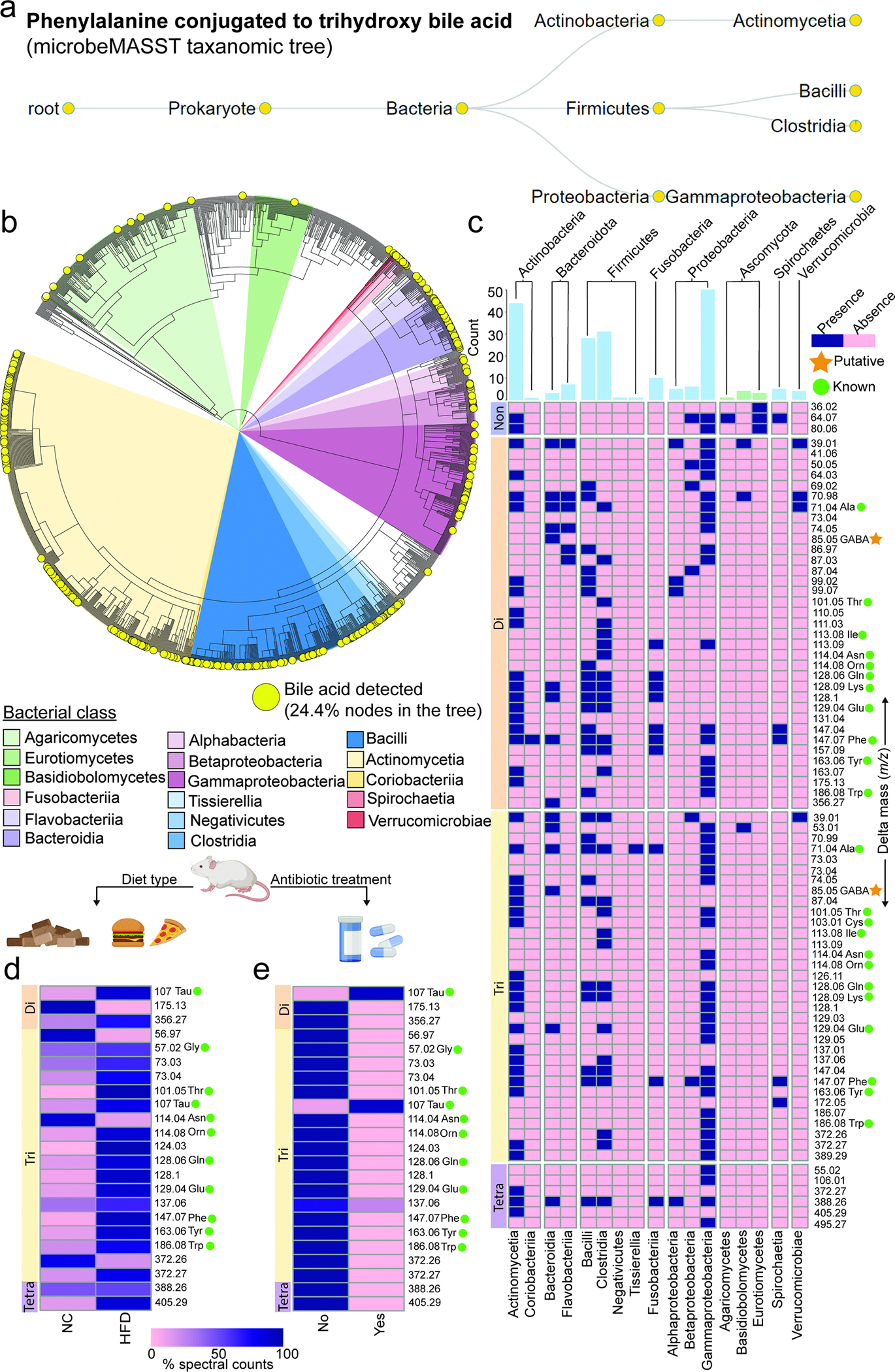
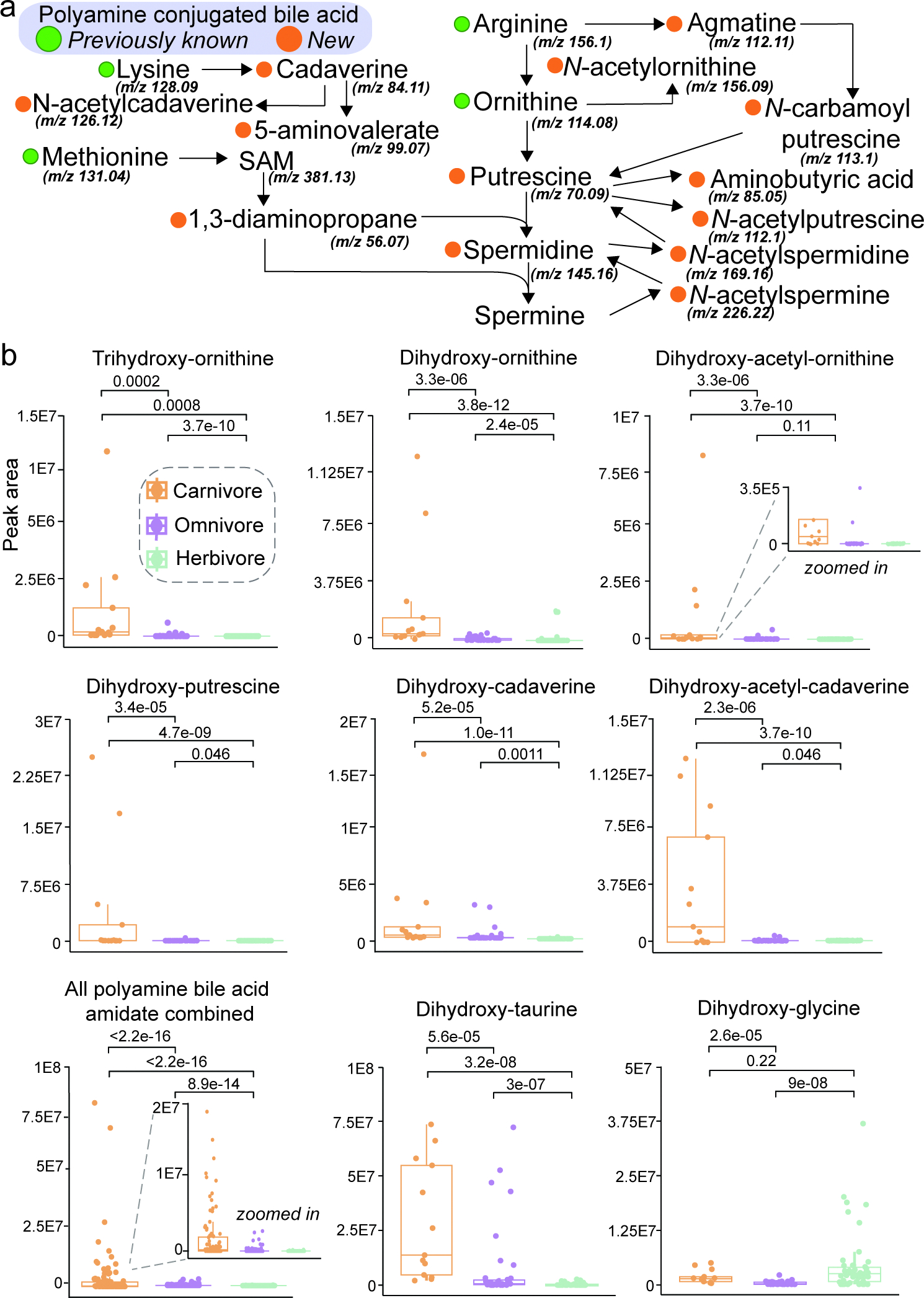
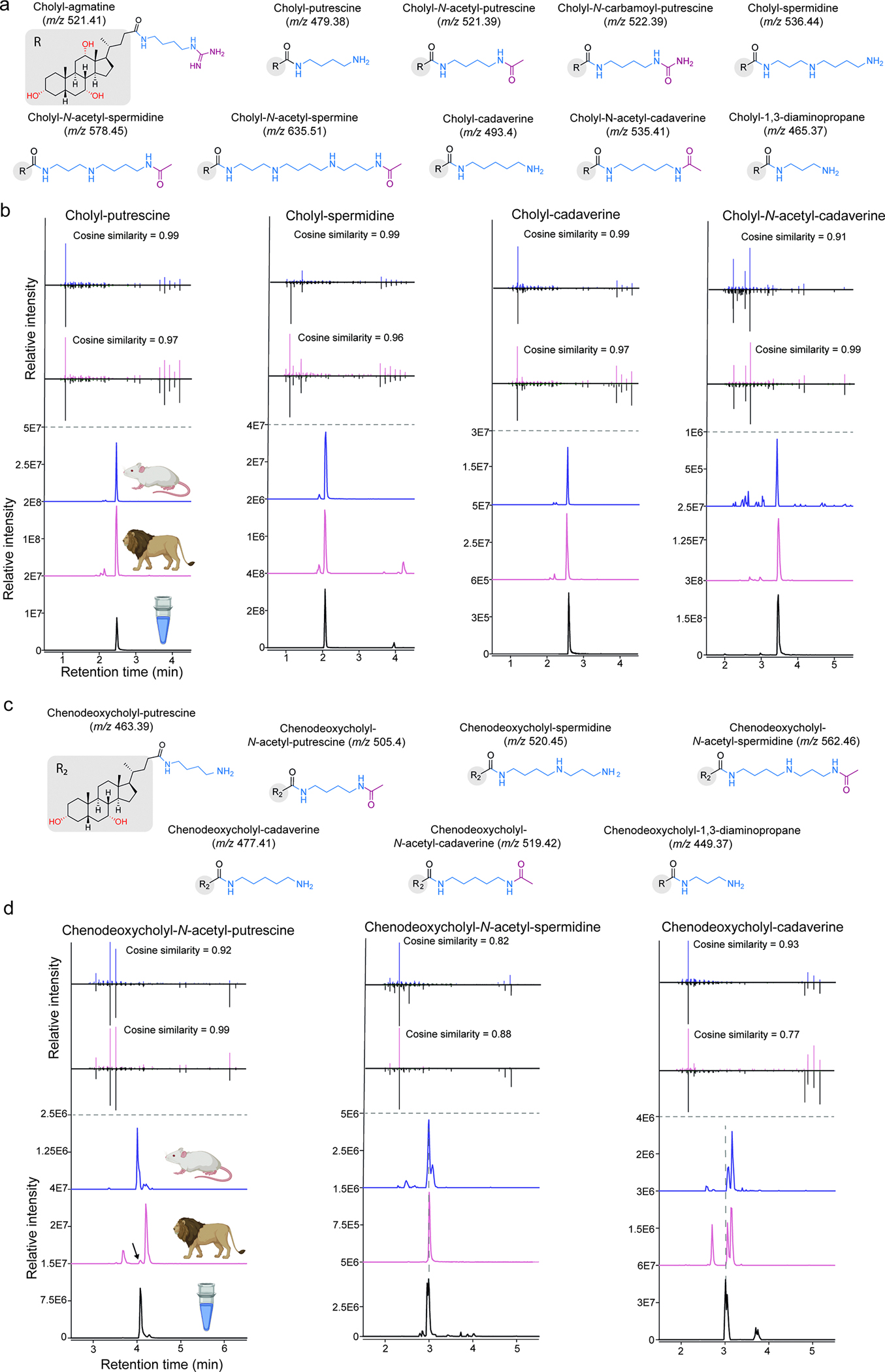
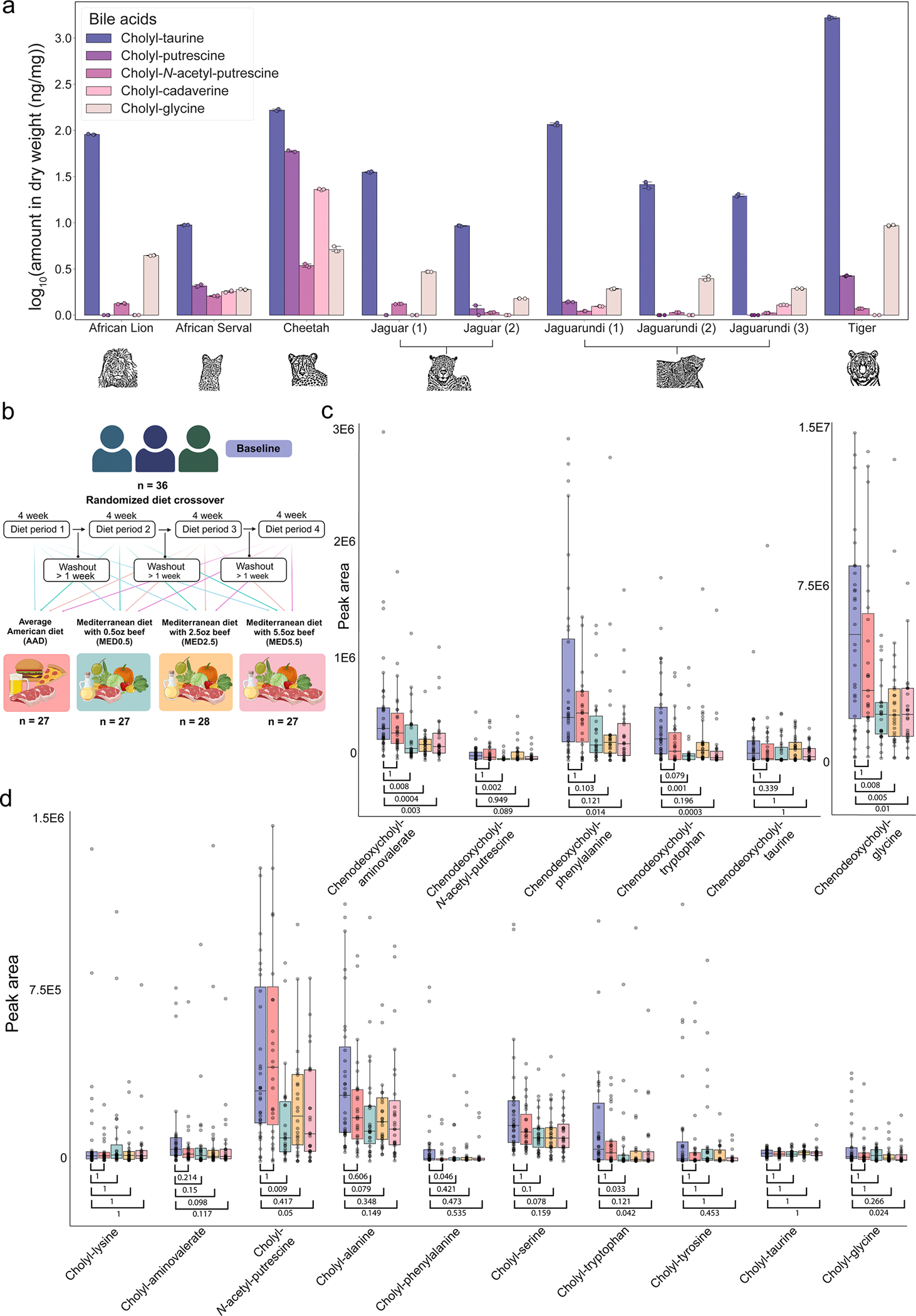
The repertoire of modifications to bile acids and related steroidal lipids by host and microbial metabolism remains incompletely characterized. To address this knowledge gap, we created a reusable resource of tandem mass spectrometry (MS/MS) spectra by filtering 1.2 billion publicly available MS/MS spectra for bile-acid-selective ion patterns. Thousands of modifications are distributed throughout animal and human bodies as well as microbial cultures. We employed this MS/MS library to identify polyamine bile amidates, prevalent in carnivores. They are present in humans, and their levels alter with a diet change from a Mediterranean to a typical American diet. This work highlights the existence of many more bile acid modifications than previously recognized and the value of leveraging public large-scale untargeted metabolomics data to discover metabolites. The availability of a modification-centric bile acid MS/MS library will inform future studies investigating bile acid roles in health and disease.
Keywords: GABA; MassQL; agmatine; bile acids; diet; fastMASST; microbial; polyamines; putrescine; spectral resource.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests P.C.D. is an advisor and holds equity in Cybele and Sirenas and a scientific co-founder, advisor, and holds equity in Ometa, Enveda, and Arome with prior approval by UC San Diego. P.C.D. also consulted for DSM animal health in 2023. M.W. is a co-founder of Ometa Labs LLC.
Figures
References
-
- Parks DJ, Blanchard SG, Bledsoe RK, Chandra G, Consler TG, Kliewer SA, Stimmel JB, Willson TM, Zavacki AM, Moore DD, et al. (1999). Bile acids: natural ligands for an orphan nuclear receptor. Science 284, 1365–1368. - PubMed
-
- Makishima M, Okamoto AY, Repa JJ, Tu H, Learned RM, Luk A, Hull MV, Lustig KD, Mangelsdorf DJ, and Shan B (1999). Identification of a nuclear receptor for bile acids. Science 284, 1362–1365. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
