

Lipid homeostasis is essential for oogenesis and embryogenesis in the silkworm, Bombyx mori
- PMID: 38472536
- PMCID: PMC10933143
- DOI: 10.1007/s00018-024-05173-8
Lipid homeostasis is essential for oogenesis and embryogenesis in the silkworm, Bombyx mori
Abstract
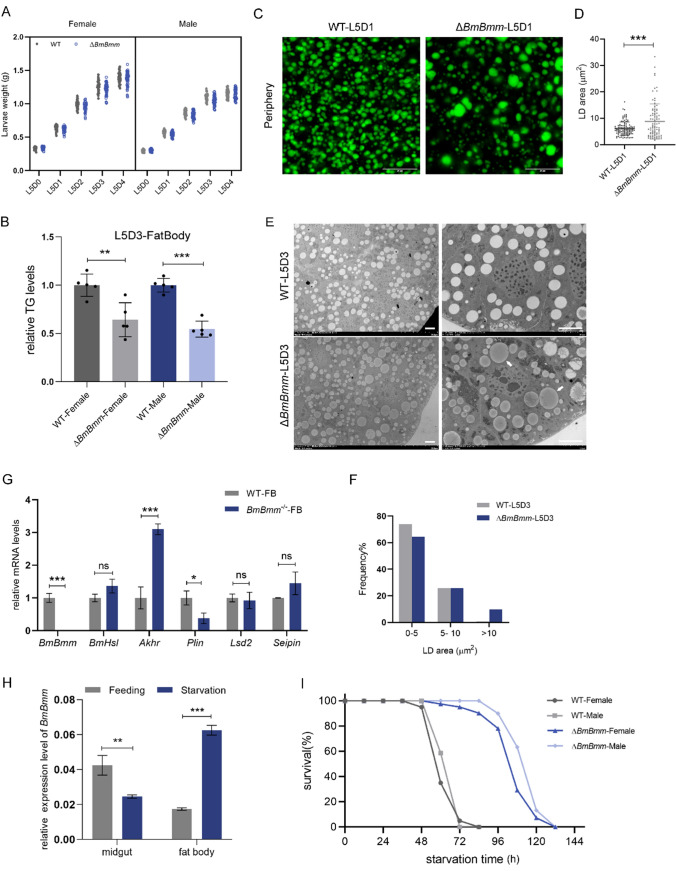
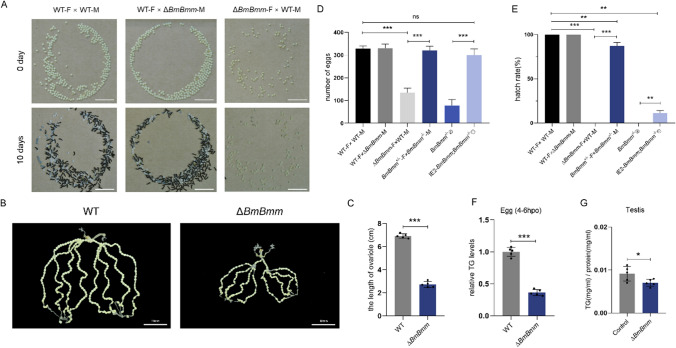
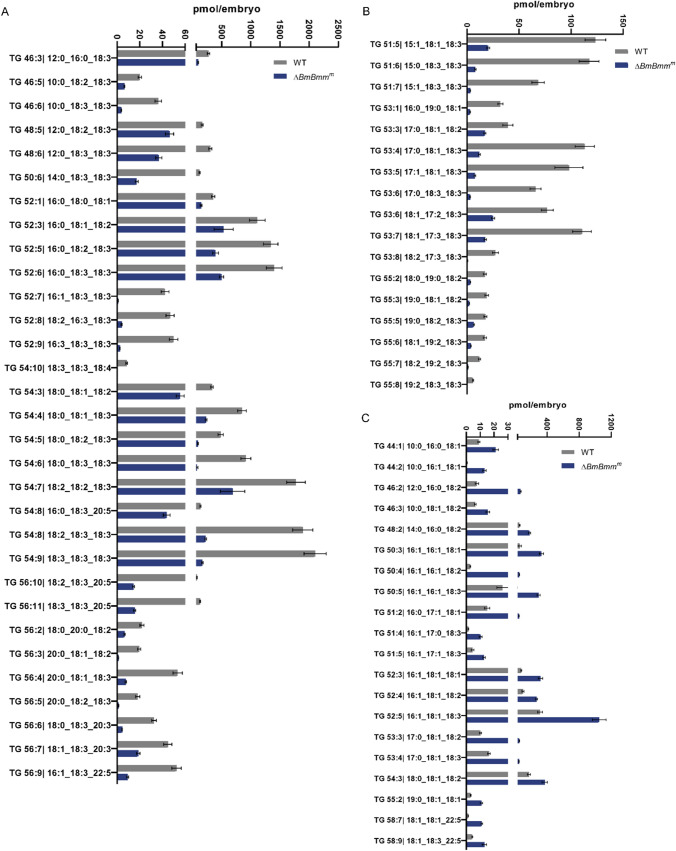
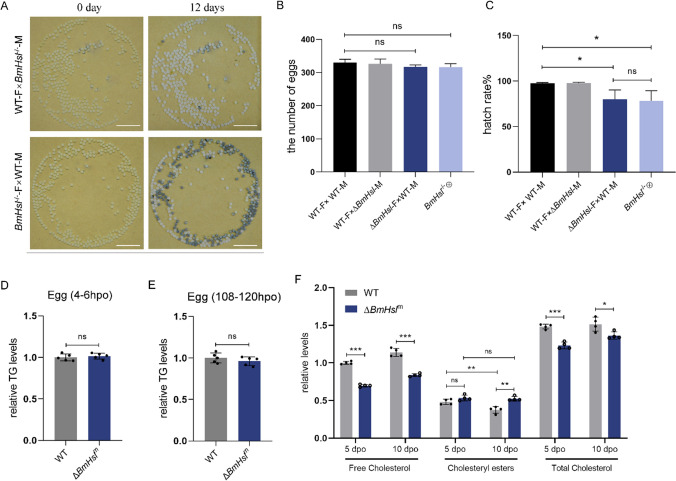
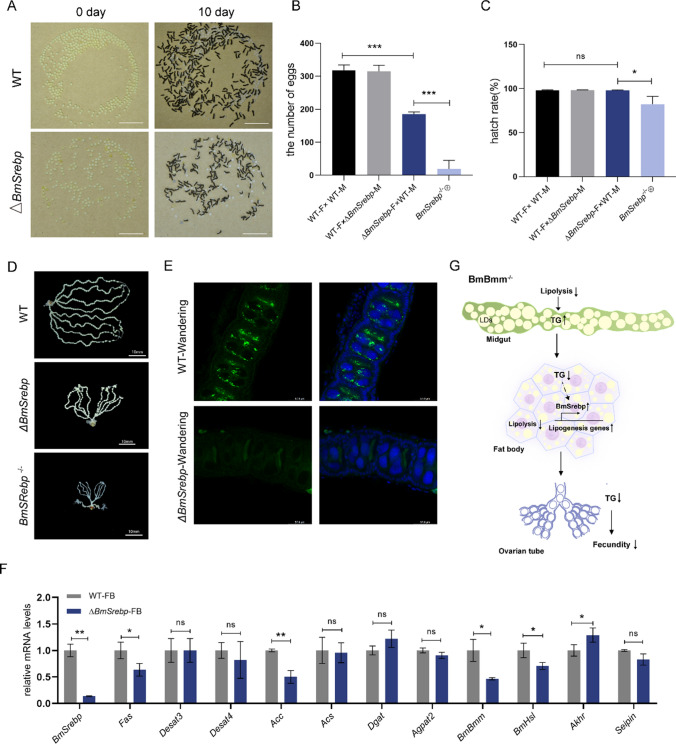
Reproduction, a fundamental feature of all known life, closely correlates with energy homeostasis. The control of synthesizing and mobilizing lipids are dynamic and well-organized processes to distribute lipid resources across tissues or generations. However, how lipid homeostasis is precisely coordinated during insect reproductive development is poorly understood. Here we describe the relations between energy metabolism and reproduction in the silkworm, Bombyx mori, a lepidopteran model insect, by using CRISPR/Cas9-mediated mutation analysis and comprehensively functional investigation on two major lipid lipases of Brummer (BmBmm) and hormone-sensitive lipase (BmHsl), and the sterol regulatory element binding protein (BmSrebp). BmBmm is a crucial regulator of lipolysis to maintain female fecundity by regulating the triglyceride (TG) storage among the midgut, the fat body, and the ovary. Lipidomics analysis reveals that defective lipolysis of females influences the composition of TG and other membrane lipids in the BmBmm mutant embryos. In contrast, BmHsl mediates embryonic development by controlling sterol metabolism rather than TG metabolism. Transcriptome analysis unveils that BmBmm deficiency significantly improves the expression of lipid synthesis-related genes including BmSrebp in the fat body. Subsequently, we identify BmSrebp as a key regulator of lipid accumulation in oocytes, which promotes oogenesis and cooperates with BmBmm to support the metabolic requirements of oocyte production. In summary, lipid homeostasis plays a vital role in supporting female reproductive success in silkworms.
Keywords: Bombyx mori; Fecundity; Lipid metabolism; Lipidomics.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


Similar articles
-
Vitellogenin receptor mutation leads to the oogenesis mutant phenotype "scanty vitellin" of the silkworm, Bombyx mori.J Biol Chem. 2013 May 10;288(19):13345-55. doi: 10.1074/jbc.M113.462556. Epub 2013 Mar 20. J Biol Chem. 2013. PMID: 23515308 Free PMC article.
-
ABHD5 regulates midgut-specific lipid homeostasis in Bombyx mori.Insect Sci. 2025 Apr;32(2):425-436. doi: 10.1111/1744-7917.13386. Epub 2024 Jun 6. Insect Sci. 2025. PMID: 38841829
-
Cyclin B3 plays pleiotropic roles in female reproductive organogenesis and early embryogenesis in the silkworm, Bombyx mori.Pest Manag Sci. 2024 Feb;80(2):376-387. doi: 10.1002/ps.7767. Epub 2023 Oct 17. Pest Manag Sci. 2024. PMID: 37698372
-
dmrt11E ortholog is a crucial factor for oogenesis of the domesticated silkworm, Bombyx mori.Insect Biochem Mol Biol. 2021 Feb;129:103517. doi: 10.1016/j.ibmb.2020.103517. Epub 2021 Jan 7. Insect Biochem Mol Biol. 2021. PMID: 33422636
-
Differentially expressed genes in the ovary of the sixth day of pupal "Ming" lethal egg mutant of silkworm, Bombyx mori.Gene. 2013 Sep 15;527(1):161-6. doi: 10.1016/j.gene.2013.05.049. Epub 2013 Jun 11. Gene. 2013. PMID: 23769927
Cited by
-
Balancing metabolism and reproduction.Elife. 2024 Apr 11;13:e97601. doi: 10.7554/eLife.97601. Elife. 2024. PMID: 38602271 Free PMC article.
-
Maternal lipid mobilization is essential for embryonic development in the malaria vector Anopheles gambiae.PLoS Biol. 2024 Dec 17;22(12):e3002960. doi: 10.1371/journal.pbio.3002960. eCollection 2024 Dec. PLoS Biol. 2024. PMID: 39689130 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
