

Myelin Basic Protein Attenuates Furin-Mediated Bri2 Cleavage and Postpones Its Membrane Trafficking
- PMID: 38473856
- PMCID: PMC10932164
- DOI: 10.3390/ijms25052608
Myelin Basic Protein Attenuates Furin-Mediated Bri2 Cleavage and Postpones Its Membrane Trafficking
Abstract
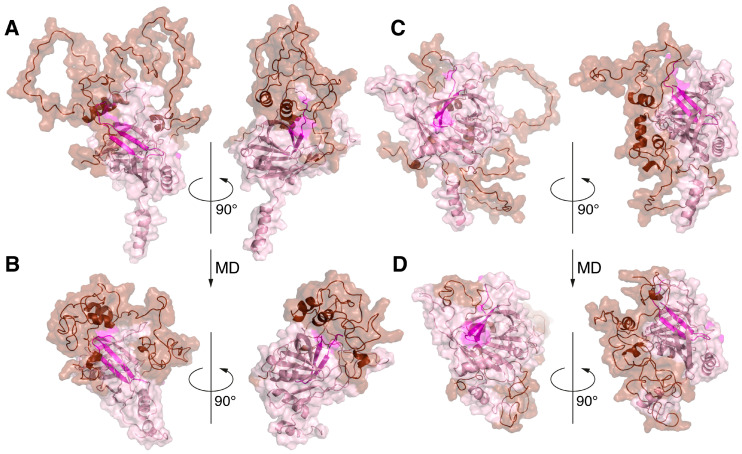
Myelin basic protein (MBP) is the second most abundant protein in the central nervous system and is responsible for structural maintenance of the myelin sheath covering axons. Previously, we showed that MBP has a more proactive role in the oligodendrocyte homeostasis, interacting with membrane-associated proteins, including integral membrane protein 2B (ITM2B or Bri2) that is associated with familial dementias. Here, we report that the molecular dynamics of the in silico-generated MBP-Bri2 complex revealed that MBP covers a significant portion of the Bri2 ectodomain, assumingly trapping the furin cleavage site, while the surface of the BRICHOS domain, which is responsible for the multimerization and activation of the Bri2 high-molecular-weight oligomer chaperone function, remains unmasked. These observations were supported by the co-expression of MBP with Bri2, its mature form, and disease-associated mutants, which showed that in mammalian cells, MBP indeed modulates the post-translational processing of Bri2 by restriction of the furin-catalyzed release of its C-terminal peptide. Moreover, we showed that the co-expression of MBP and Bri2 also leads to an altered cellular localization of Bri2, restricting its membrane trafficking independently of the MBP-mediated suppression of the Bri2 C-terminal peptide release. Further investigations should elucidate if these observations have physiological meaning in terms of Bri2 as a MBP chaperone activated by the MBP-dependent postponement of Bri2 membrane trafficking.
Keywords: artificial intelligence; integral membrane protein 2B (ITM2B/Bri2); intermolecular interaction; myelin basic protein (MBP); protein folding; protein processing regulation.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures








Similar articles
-
Glycosylation of BRI2 on asparagine 170 is involved in its trafficking to the cell surface but not in its processing by furin or ADAM10.Glycobiology. 2011 Oct;21(10):1382-8. doi: 10.1093/glycob/cwr097. Epub 2011 Jul 13. Glycobiology. 2011. PMID: 21752865 Free PMC article.
-
Regulated intramembrane proteolysis of Bri2 (Itm2b) by ADAM10 and SPPL2a/SPPL2b.J Biol Chem. 2008 Jan 18;283(3):1644-1652. doi: 10.1074/jbc.M706661200. Epub 2007 Oct 25. J Biol Chem. 2008. PMID: 17965014
-
Identification of glycolytic proteins as binding partners of Bri2 BRICHOS domain.J Pharm Biomed Anal. 2023 Aug 5;232:115465. doi: 10.1016/j.jpba.2023.115465. Epub 2023 May 18. J Pharm Biomed Anal. 2023. PMID: 37220701 Free PMC article.
-
Role of BRI2 in dementia.J Alzheimers Dis. 2014;40(3):481-94. doi: 10.3233/JAD-131364. J Alzheimers Dis. 2014. PMID: 24473189 Review.
-
BRI2 as a central protein involved in neurodegeneration.Biotechnol J. 2008 Dec;3(12):1548-54. doi: 10.1002/biot.200800247. Biotechnol J. 2008. PMID: 19072909 Review.
References
-
- Benson M.D., Buxbaum J.N., Eisenberg D.S., Merlini G., Saraiva M.J.M., Sekijima Y., Sipe J.D., Westermark P. Amyloid Nomenclature 2018: Recommendations by the International Society of Amyloidosis (ISA) Nomenclature Committee. Amyloid. 2018;25:215–219. doi: 10.1080/13506129.2018.1549825. - DOI - PubMed
-
- Alzheimer Disease Genetics Consortium (ADGC) The European Alzheimer’s Disease Initiative (EADI) Cohorts for Heart and Aging Research in Genomic Epidemiology Consortium (CHARGE) Genetic and Environmental Risk in AD/Defining Genetic, Polygenic and Environmental Risk for Alzheimer’s Disease Consortium (GERAD/PERADES) Kunkle B.W., Grenier-Boley B., Sims R., Bis J.C., Damotte V., Naj A.C., et al. Genetic Meta-Analysis of Diagnosed Alzheimer’s Disease Identifies New Risk Loci and Implicates Aβ, Tau, Immunity and Lipid Processing. Nat. Genet. 2019;51:414–430. doi: 10.1038/s41588-019-0358-2. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
