

Evaluating Temperature Effects on Bluetongue Virus Serotype 10 and 17 Coinfection in Culicoides sonorensis
- PMID: 38474308
- PMCID: PMC10932384
- DOI: 10.3390/ijms25053063
Evaluating Temperature Effects on Bluetongue Virus Serotype 10 and 17 Coinfection in Culicoides sonorensis
Abstract
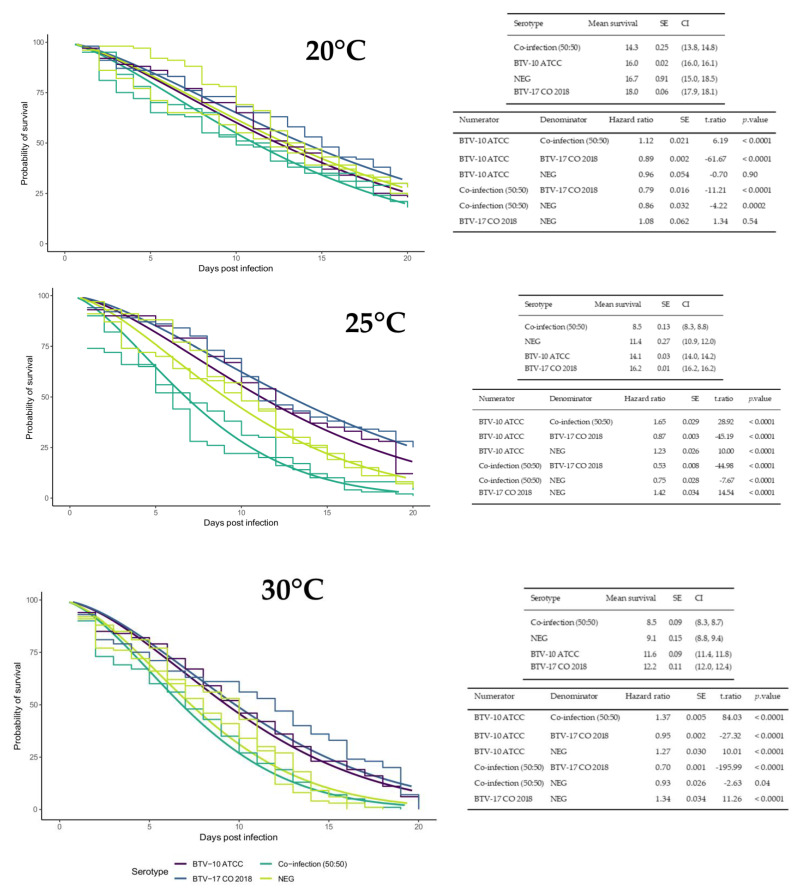
Bluetongue virus (BTV) is a segmented, double-stranded RNA virus transmitted by Culicoides midges that infects ruminants. As global temperatures increase and geographical ranges of midges expand, there is increased potential for BTV outbreaks from incursions of novel serotypes into endemic regions. However, an understanding of the effect of temperature on reassortment is lacking. The objectives of this study were to compare how temperature affected Culicoides survival, virogenesis, and reassortment in Culicoides sonorensis coinfected with two BTV serotypes. Midges were fed blood meals containing BTV-10, BTV-17, or BTV serotype 10 and 17 and maintained at 20 °C, 25 °C, or 30 °C. Midge survival was assessed, and pools of midges were collected every other day to evaluate virogenesis of BTV via qRT-PCR. Additional pools of coinfected midges were collected for BTV plaque isolation. The genotypes of plaques were determined using next-generation sequencing. Warmer temperatures impacted traits related to vector competence in offsetting ways: BTV replicated faster in midges at warmer temperatures, but midges did not survive as long. Overall, plaques with BTV-17 genotype dominated, but BTV-10 was detected in some plaques, suggesting parental strain fitness may play a role in reassortment outcomes. Temperature adds an important dimension to host-pathogen interactions with implications for transmission and evolution.
Keywords: Culicoides; bluetongue virus; co-infection; next-generation sequencing; reassortment; temperature.
Conflict of interest statement
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Figures
References
-
- Navarro Mamani D.A., Ramos Huere H., Vera Buendia R., Rojas M., Chunga W.A., Valdez Gutierrez E., Vergara Abarca W., Rivera Gerónimo H., Altamiranda-Saavedra M. Would Climate Change Influence the Potential Distribution and Ecological Niche of Bluetongue Virus and Its Main Vector in Peru? Viruses. 2023;15:892. doi: 10.3390/v15040892. - DOI - PMC - PubMed
-
- Möhlmann T.W.R., Keeling M.J., Wennergren U., Favia G., Santman-Berends I., Takken W., Koenraadt C.J.M., Brand S.P.C. Biting Midge Dynamics and Bluetongue Transmission: A Multiscale Model Linking Catch Data with Climate and Disease Outbreaks. Sci. Rep. 2021;11:1892. doi: 10.1038/s41598-021-81096-9. - DOI - PMC - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
