

Mechanoecology: biomechanical aspects of insect-plant interactions
- PMID: 38480551
- PMCID: PMC10994878
- DOI: 10.1007/s00359-024-01698-2
Mechanoecology: biomechanical aspects of insect-plant interactions
Abstract
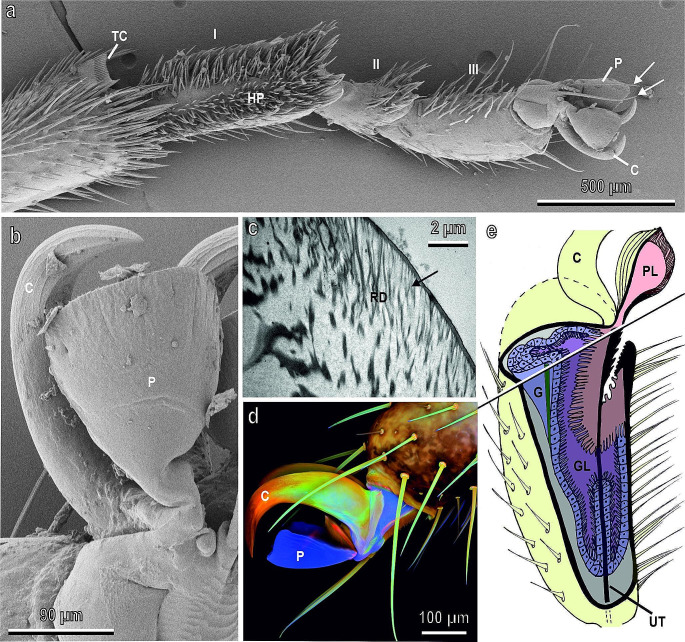
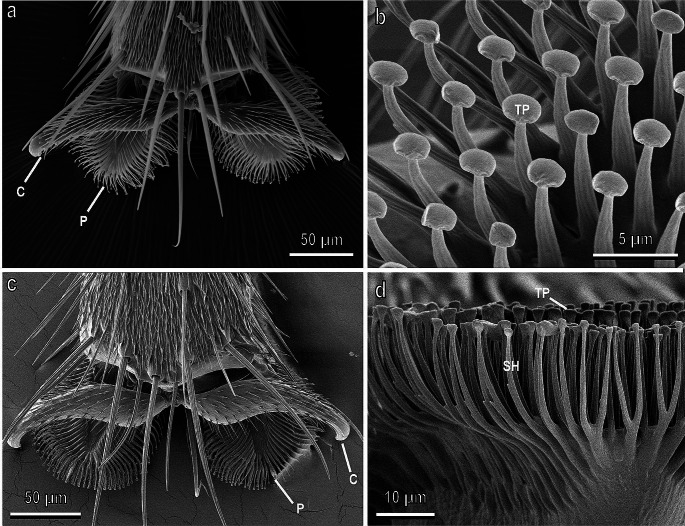
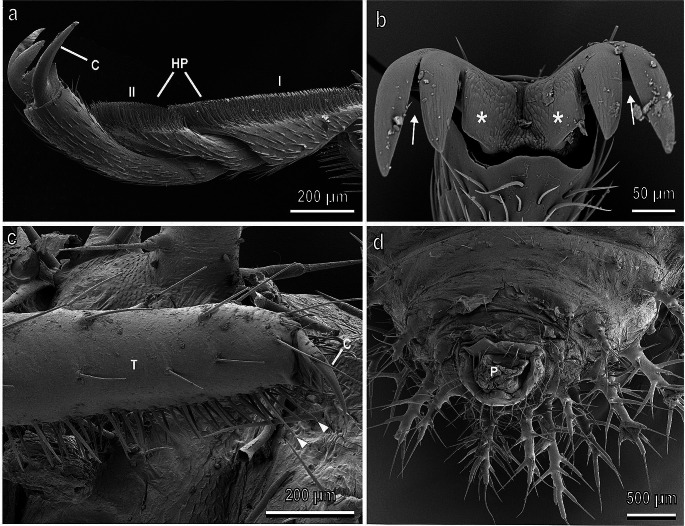
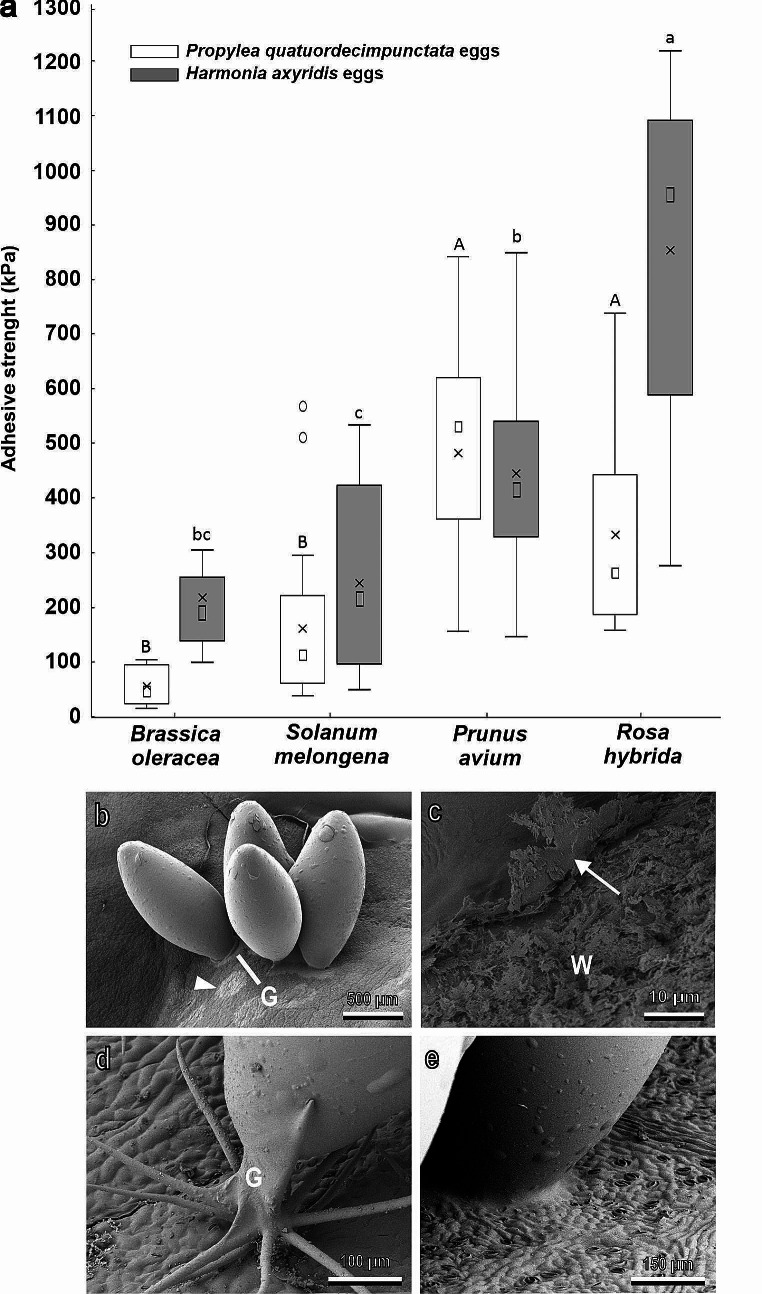
Plants and herbivorous insects as well as their natural enemies, such as predatory and parasitoid insects, are united by intricate relationships. During the long period of co-evolution with insects, plants developed a wide diversity of features to defence against herbivores and to attract pollinators and herbivores' natural enemies. The chemical basis of insect-plant interactions is established and many examples are studied, where feeding and oviposition site selection of phytophagous insects are dependent on the plant's secondary chemistry. However, often overlooked mechanical interactions between insects and plants can be rather crucial. In the context of mechanoecology, the evolution of plant surfaces and insect adhesive pads is an interesting example of competition between insect attachment systems and plant anti-attachment surfaces. The present review is focused on mechanical insect-plant interactions of some important pest species, such as the polyphagous Southern Green Stinkbug Nezara viridula and two frugivorous pest species, the polyphagous Mediterranean fruit fly Ceratitis capitata and the monophagous olive fruit fly Bactrocera oleae. Their ability to attach to plant surfaces characterised by different features such as waxes and trichomes is discussed. Some attention is paid also to Coccinellidae, whose interaction with plant leaf surfaces is substantial across all developmental stages in both phytophagous and predatory species that feed on herbivorous insects. Finally, the role of different kinds of anti-adhesive nanomaterials is discussed. They can reduce the attachment ability of insect pests to natural and artificial surfaces, potentially representing environmental friendly alternative methods to reduce insect pest impact in agriculture.
Keywords: Adhesion; Adhesive pad; Biomechanics; Ecology; Epicuticular wax; Friction; Insects; Plants; Surface.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing or financial interests.
Figures


References
-
- Akandeh M, Shishehbor P. Life history traits of melon ladybeetle, Epilachna Chrysomelina (Col.: Coccinellidae), on four host plant species. J Entomol Soc Iran. 2011;31:17–27.
-
- Al Bitar L, Gorb SN, Zebitz CPW, Voigt D. Egg adhesion of the codling moth Cydia Pomonella L. (Lepidoptera, Tortricidae) to various substrates: II. Fruit surfaces of different apple cultivars. Arthropod Plant Interact. 2014;8:57–77. doi: 10.1007/s11829-013-9288-6. - DOI
-
- Atkin DSJ, Hamilton RJ. The effects of plant waxes on insects. J Nat Prod. 1982;45:694–696. doi: 10.1021/np50024a007. - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
