

IL-6 signaling drives self-renewal and alternative activation of adipose tissue macrophages
- PMID: 38482013
- PMCID: PMC10933059
- DOI: 10.3389/fimmu.2024.1201439
IL-6 signaling drives self-renewal and alternative activation of adipose tissue macrophages
Abstract
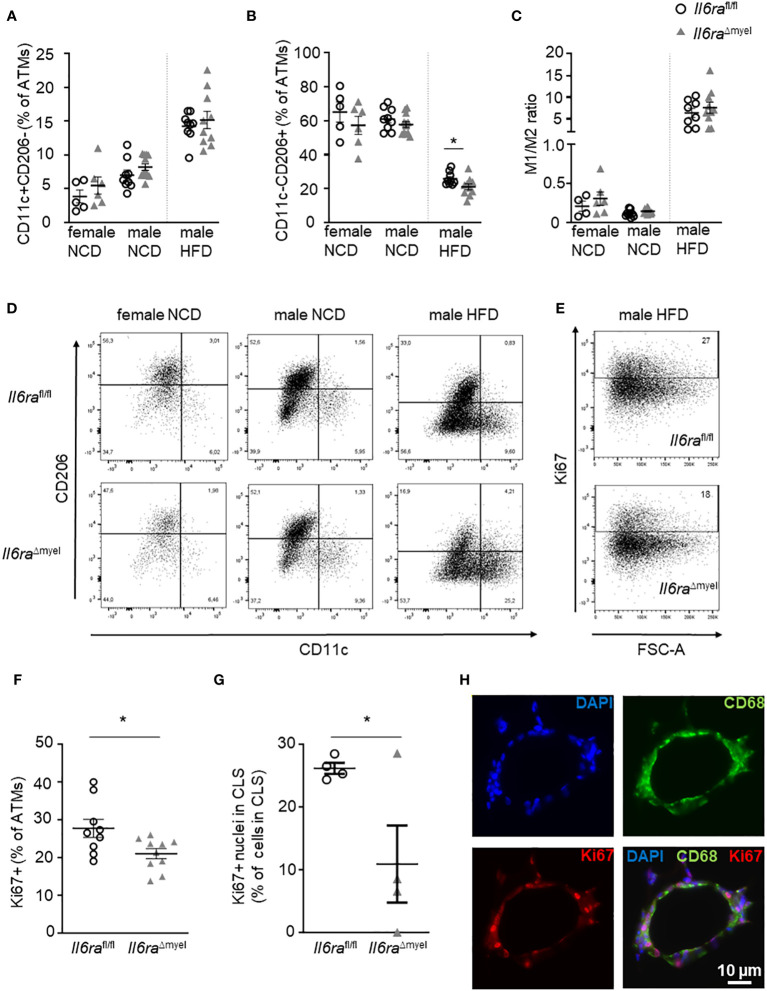
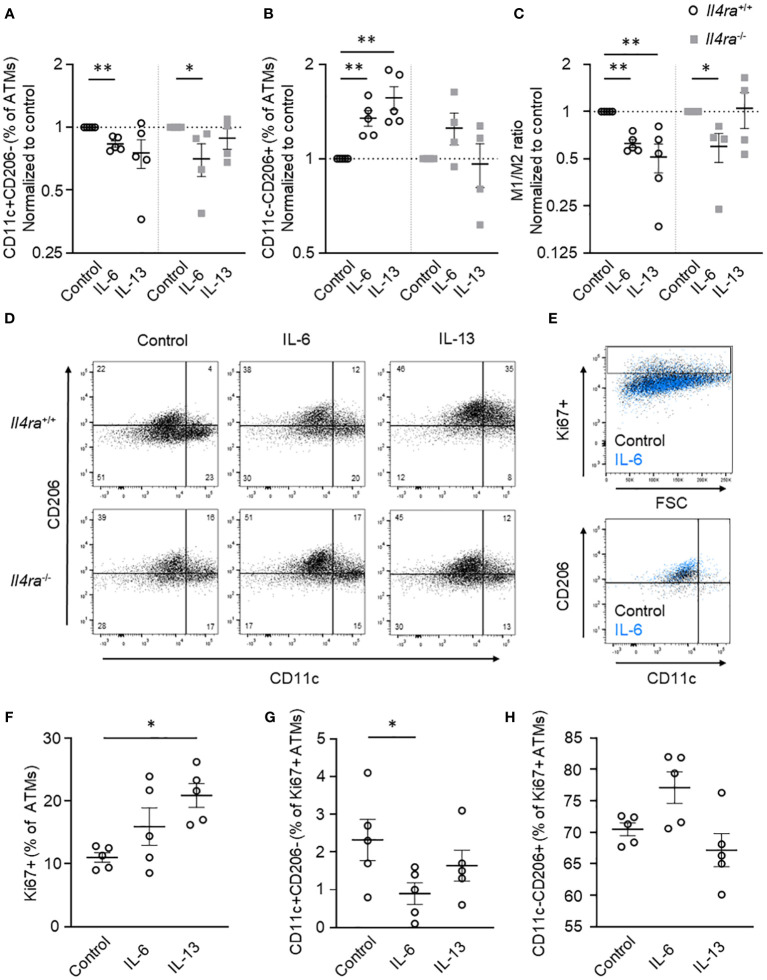
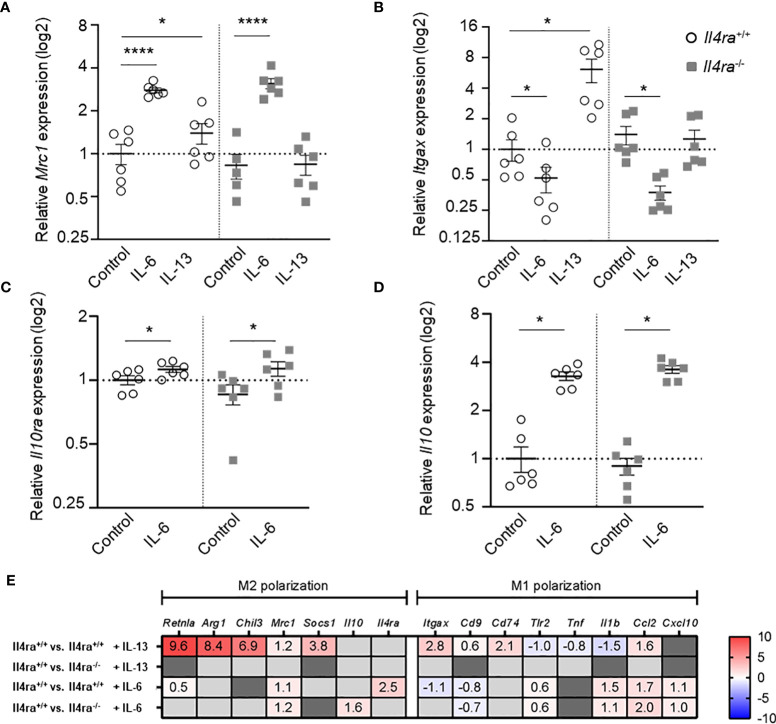
Introduction: Obesity is associated with chronic low-grade inflammation of adipose tissue (AT) and an increase of AT macrophages (ATMs) that is linked to the onset of type 2 diabetes. We have recently shown that neutralization of interleukin (IL)-6 in obese AT organ cultures inhibits proliferation of ATMs, which occurs preferentially in alternatively activated macrophage phenotype.
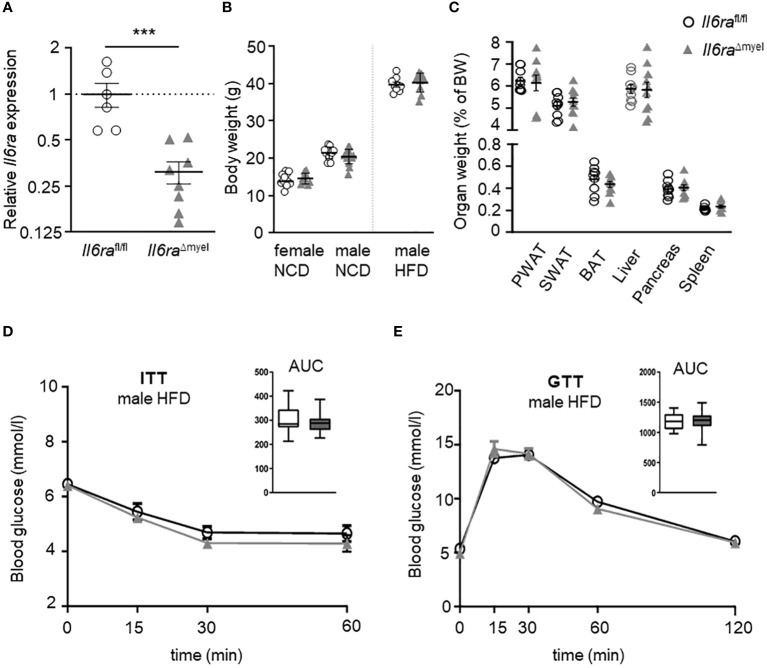
Methods: In this study, we investigated AT biology and the metabolic phenotype of mice with myeloid cell-specific IL-6Rα deficiency (Il6ra Δmyel) after normal chow and 20 weeks of high-fat diet focusing on AT inflammation, ATM polarization and proliferation. Using organotypical AT culture and bone marrow derived macrophages (BMDMs) of IL-4Rα knockout mice (Il4ra -/-) we studied IL-6 signaling.
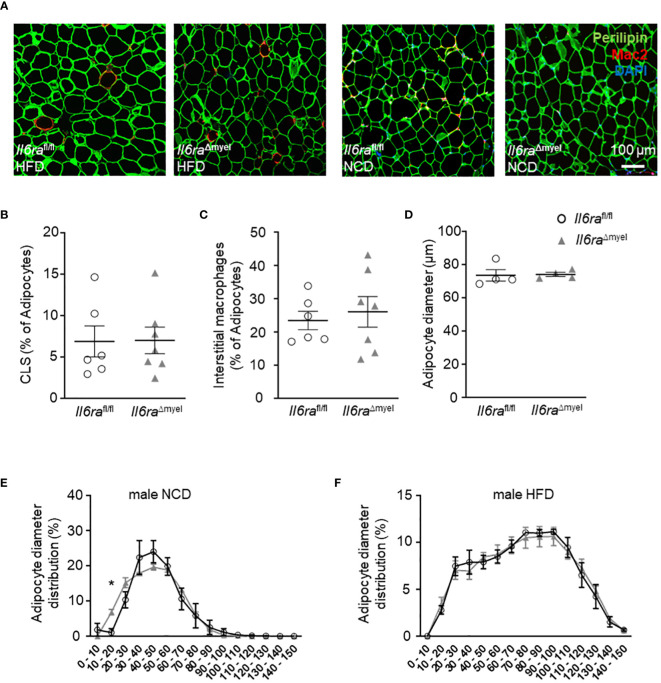
Results: Obese Il6ra Δmyel mice exhibited no differences in insulin sensitivity or histological markers of AT inflammation. Notably, we found a reduction of ATMs expressing the mannose receptor 1 (CD206), as well as a decrease of the proliferation marker Ki67 in ATMs of Il6ra Δmyel mice. Importantly, organotypical AT culture and BMDM data of Il4ra -/- mice revealed that IL-6 mediates a shift towards the M2 phenotype independent from the IL-6/IL-4Rα axis.
Discussion: Our results demonstrate IL-4Rα-independent anti-inflammatory effects of IL-6 on macrophages and the ability of IL-6 to maintain proliferation rates in obese AT.
Keywords: IL-6; adipose tissue inflammation; alternative activation; diabetes; macrophages; mannose receptor; obesity; self-renewal.
Copyright © 2024 Ackermann, Arndt, Fröba, Lindhorst, Glaß, Kirstein, Hobusch, Wunderlich, Braune and Gericke.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
Myeloid Cell-Specific IL-4 Receptor Knockout Partially Protects from Adipose Tissue Inflammation.J Immunol. 2021 Dec 15;207(12):3081-3089. doi: 10.4049/jimmunol.2100699. Epub 2021 Nov 17. J Immunol. 2021. PMID: 34789558
-
Local proliferation of macrophages in adipose tissue during obesity-induced inflammation.Diabetologia. 2014 Mar;57(3):562-71. doi: 10.1007/s00125-013-3139-y. Epub 2013 Dec 17. Diabetologia. 2014. PMID: 24343232
-
MicroRNA-30 modulates metabolic inflammation by regulating Notch signaling in adipose tissue macrophages.Int J Obes (Lond). 2018 Jun;42(6):1140-1150. doi: 10.1038/s41366-018-0114-1. Epub 2018 Jun 13. Int J Obes (Lond). 2018. PMID: 29899524 Free PMC article.
-
Adipose tissue macrophages.Immunol Lett. 2007 Oct 15;112(2):61-7. doi: 10.1016/j.imlet.2007.07.003. Epub 2007 Jul 31. Immunol Lett. 2007. PMID: 17719095 Review.
-
The "Big Bang" in obese fat: Events initiating obesity-induced adipose tissue inflammation.Eur J Immunol. 2015 Sep;45(9):2446-56. doi: 10.1002/eji.201545502. Epub 2015 Aug 19. Eur J Immunol. 2015. PMID: 26220361 Review.
Cited by
-
AhR governs lipid metabolism: the role of gut microbiota.Front Microbiol. 2025 Jan 29;16:1442282. doi: 10.3389/fmicb.2025.1442282. eCollection 2025. Front Microbiol. 2025. PMID: 39944639 Free PMC article. Review.
-
The multifaceted regulation of white adipose tissue browning and their therapeutic potential.J Physiol Biochem. 2025 Aug 11. doi: 10.1007/s13105-025-01117-3. Online ahead of print. J Physiol Biochem. 2025. PMID: 40785003 Review.
-
Effects of Bariatric Surgery-Related Weight Loss on the Characteristics, Metabolism, and Immunomodulation of Adipose Stromal/Stem Cells in a Follow-Up Study.Stem Cells Int. 2025 May 13;2025:1212255. doi: 10.1155/sci/1212255. eCollection 2025. Stem Cells Int. 2025. PMID: 40395977 Free PMC article.
-
Lymphocytes and monocytes undergo swift suppression of IL-10R, IL-6R, and IL-2Rβγ signaling under high concentrations of different cytokines.bioRxiv [Preprint]. 2025 May 8:2025.05.02.651967. doi: 10.1101/2025.05.02.651967. bioRxiv. 2025. PMID: 40654786 Free PMC article. Preprint.
-
Trends and Hot Spots of Macrophages Linked to Metabolic Syndrome: A Comprehensive Bibliometric and Visualization Analysis (2014-2024).Mediators Inflamm. 2025 May 24;2025:9487093. doi: 10.1155/mi/9487093. eCollection 2025. Mediators Inflamm. 2025. PMID: 40453651 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous