

JUN mRNA translation regulation is mediated by multiple 5' UTR and start codon features
- PMID: 38483896
- PMCID: PMC10939236
- DOI: 10.1371/journal.pone.0299779
JUN mRNA translation regulation is mediated by multiple 5' UTR and start codon features
Abstract
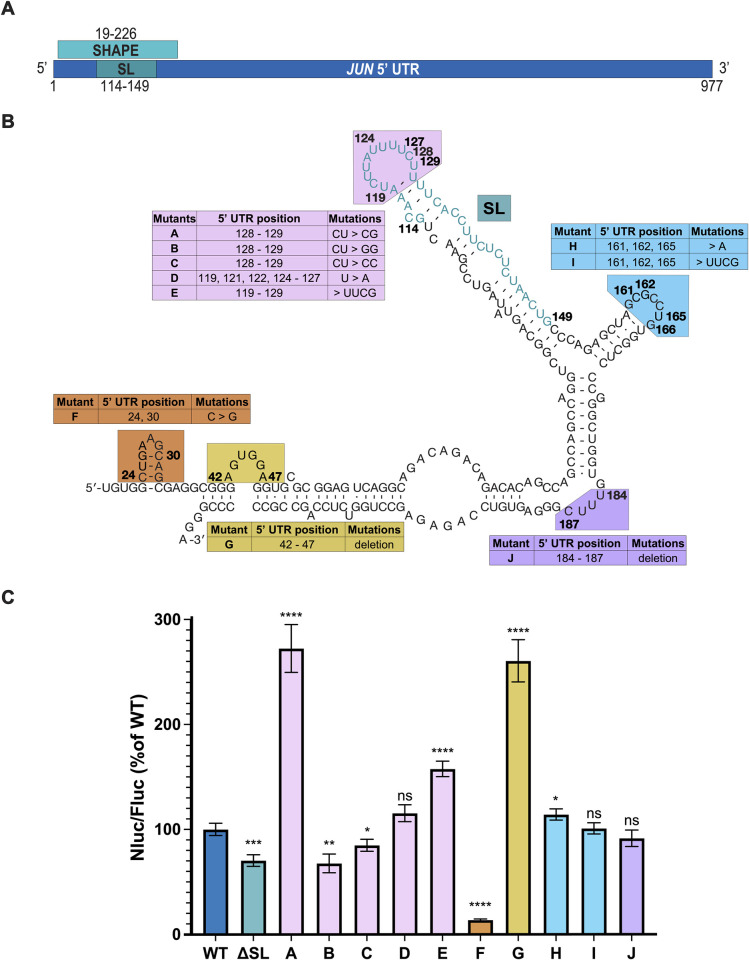
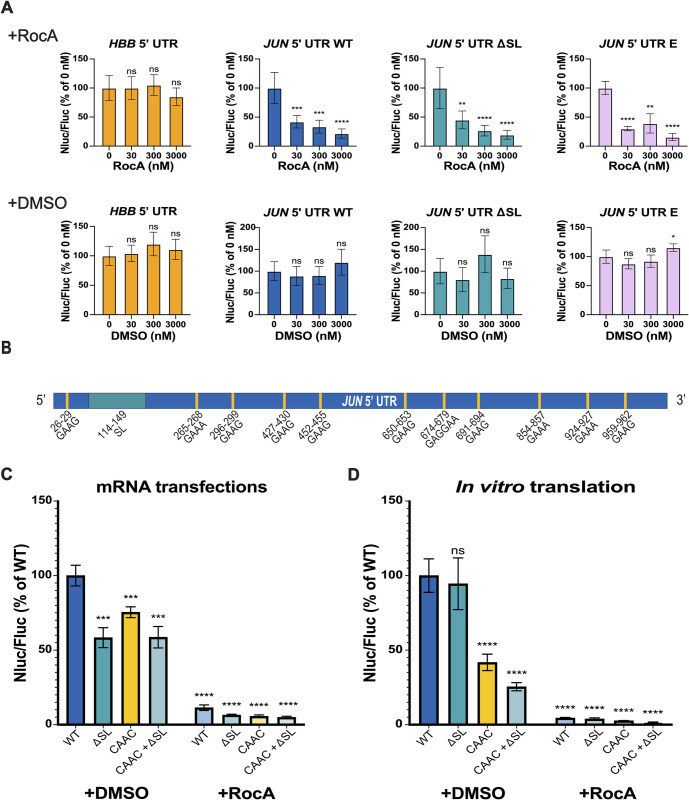
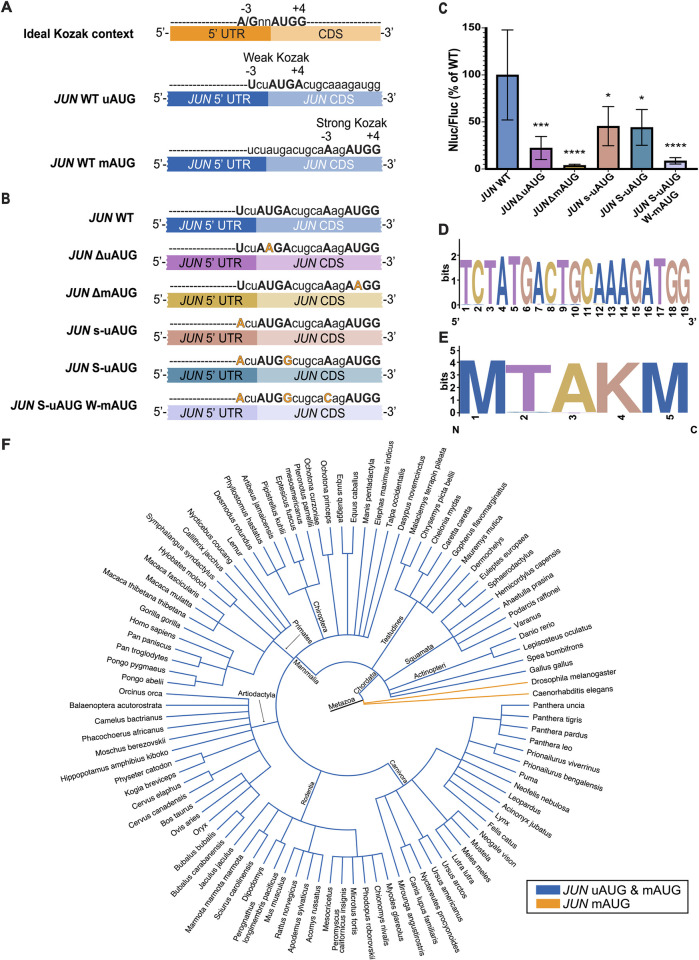
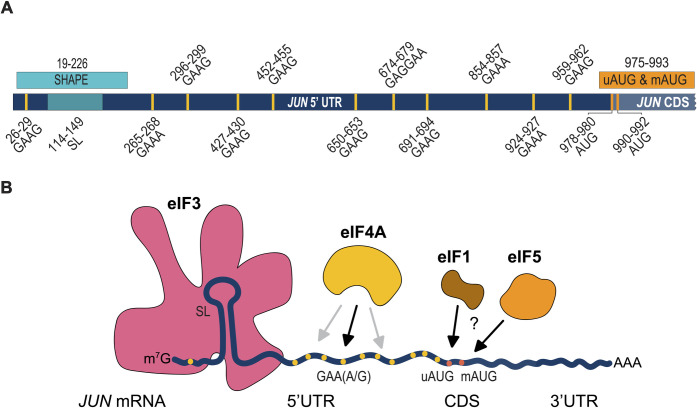
Regulation of mRNA translation by eukaryotic initiation factors (eIFs) is crucial for cell survival. In humans, eIF3 stimulates translation of the JUN mRNA which encodes the transcription factor JUN, an oncogenic transcription factor involved in cell cycle progression, apoptosis, and cell proliferation. Previous studies revealed that eIF3 activates translation of the JUN mRNA by interacting with a stem loop in the 5' untranslated region (5' UTR) and with the 5' -7-methylguanosine cap structure. In addition to its interaction site with eIF3, the JUN 5' UTR is nearly one kilobase in length, and has a high degree of secondary structure, high GC content, and an upstream start codon (uAUG). This motivated us to explore the complexity of JUN mRNA translation regulation in human cells. Here we find that JUN translation is regulated in a sequence and structure-dependent manner in regions adjacent to the eIF3-interacting site in the JUN 5' UTR. Furthermore, we identify contributions of an additional initiation factor, eIF4A, in JUN regulation. We show that enhancing the interaction of eIF4A with JUN by using the compound Rocaglamide A (RocA) represses JUN translation. We also find that both the upstream AUG (uAUG) and the main AUG (mAUG) contribute to JUN translation and that they are conserved throughout vertebrates. Our results reveal additional layers of regulation for JUN translation and show the potential of JUN as a model transcript for understanding multiple interacting modes of translation regulation.
Copyright: © 2024 González-Sánchez et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Update of
-
JUN mRNA Translation Regulation is Mediated by Multiple 5' UTR and Start Codon Features.bioRxiv [Preprint]. 2023 Nov 17:2023.11.17.567602. doi: 10.1101/2023.11.17.567602. bioRxiv. 2023. Update in: PLoS One. 2024 Mar 14;19(3):e0299779. doi: 10.1371/journal.pone.0299779. PMID: 38014201 Free PMC article. Updated. Preprint.
Similar articles
-
JUN mRNA Translation Regulation is Mediated by Multiple 5' UTR and Start Codon Features.bioRxiv [Preprint]. 2023 Nov 17:2023.11.17.567602. doi: 10.1101/2023.11.17.567602. bioRxiv. 2023. Update in: PLoS One. 2024 Mar 14;19(3):e0299779. doi: 10.1371/journal.pone.0299779. PMID: 38014201 Free PMC article. Updated. Preprint.
-
Evidence That Base-pairing Interaction between Intron and mRNA Leader Sequences Inhibits Initiation of HAC1 mRNA Translation in Yeast.J Biol Chem. 2015 Sep 4;290(36):21821-32. doi: 10.1074/jbc.M115.649335. Epub 2015 Jul 14. J Biol Chem. 2015. PMID: 26175153 Free PMC article.
-
Structure of the RNA Specialized Translation Initiation Element that Recruits eIF3 to the 5'-UTR of c-Jun.J Mol Biol. 2020 Mar 27;432(7):1841-1855. doi: 10.1016/j.jmb.2020.01.001. Epub 2020 Jan 14. J Mol Biol. 2020. PMID: 31953146 Free PMC article.
-
Hepatitis C Virus Translation Regulation.Int J Mol Sci. 2020 Mar 27;21(7):2328. doi: 10.3390/ijms21072328. Int J Mol Sci. 2020. PMID: 32230899 Free PMC article. Review.
-
Human eIF3: from 'blobology' to biological insight.Philos Trans R Soc Lond B Biol Sci. 2017 Mar 19;372(1716):20160176. doi: 10.1098/rstb.2016.0176. Philos Trans R Soc Lond B Biol Sci. 2017. PMID: 28138064 Free PMC article. Review.
Cited by
-
Decoding post-transcriptional regulatory networks by RNA-linked CRISPR screening in human cells.Nat Methods. 2025 Jun;22(6):1237-1246. doi: 10.1038/s41592-025-02702-6. Epub 2025 May 29. Nat Methods. 2025. PMID: 40442371
-
Anti-tumor effects of the eIF4A inhibitor didesmethylrocaglamide and its derivatives in human and canine osteosarcomas.Sci Rep. 2024 Aug 20;14(1):19349. doi: 10.1038/s41598-024-69171-3. Sci Rep. 2024. PMID: 39164287 Free PMC article.
-
Distinct non-canonical translation initiation modes arise for specific host and viral mRNAs during poxvirus-induced shutoff.Nat Microbiol. 2025 Jun;10(6):1535-1549. doi: 10.1038/s41564-025-02009-4. Epub 2025 May 28. Nat Microbiol. 2025. PMID: 40437296
-
Anti-tumor Effects of the eIF4A Inhibitor Didesmethylrocaglamide and Its Derivatives in Human and Canine Osteosarcomas.Res Sq [Preprint]. 2024 Jun 14:rs.3.rs-4494024. doi: 10.21203/rs.3.rs-4494024/v1. Res Sq. 2024. Update in: Sci Rep. 2024 Aug 20;14(1):19349. doi: 10.1038/s41598-024-69171-3. PMID: 38947012 Free PMC article. Updated. Preprint.
-
A dynamic compositional equilibrium governs mRNA recognition by eIF3.bioRxiv [Preprint]. 2024 Aug 18:2024.04.25.581977. doi: 10.1101/2024.04.25.581977. bioRxiv. 2024. PMID: 38712078 Free PMC article. Preprint.
References
-
- Buttgereit F, Brand MD. A hierarchy of ATP-consuming processes in mammalian cells. Biochem J. 1995;312(Pt 1):163–7. Available from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1136240/ doi: 10.1042/bj3120163 - DOI - PMC - PubMed
-
- Sonenberg N, Hinnebusch AG. Regulation of Translation Initiation in Eukaryotes: Mechanisms and Biological Targets. Cell. 2009;136(4):731–45. Available from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3610329/ doi: 10.1016/j.cell.2009.01.042 - DOI - PMC - PubMed
-
- Leibovitch M, Topisirovic I. Dysregulation of mRNA translation and energy metabolism in cancer. Adv Biol Regul. 2018;67:30–9. Available from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5993950/ doi: 10.1016/j.jbior.2017.11.001 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
