

Identification of a negative-strand RNA virus with natural plant and fungal hosts
- PMID: 38483998
- PMCID: PMC10962957
- DOI: 10.1073/pnas.2319582121
Identification of a negative-strand RNA virus with natural plant and fungal hosts
Abstract
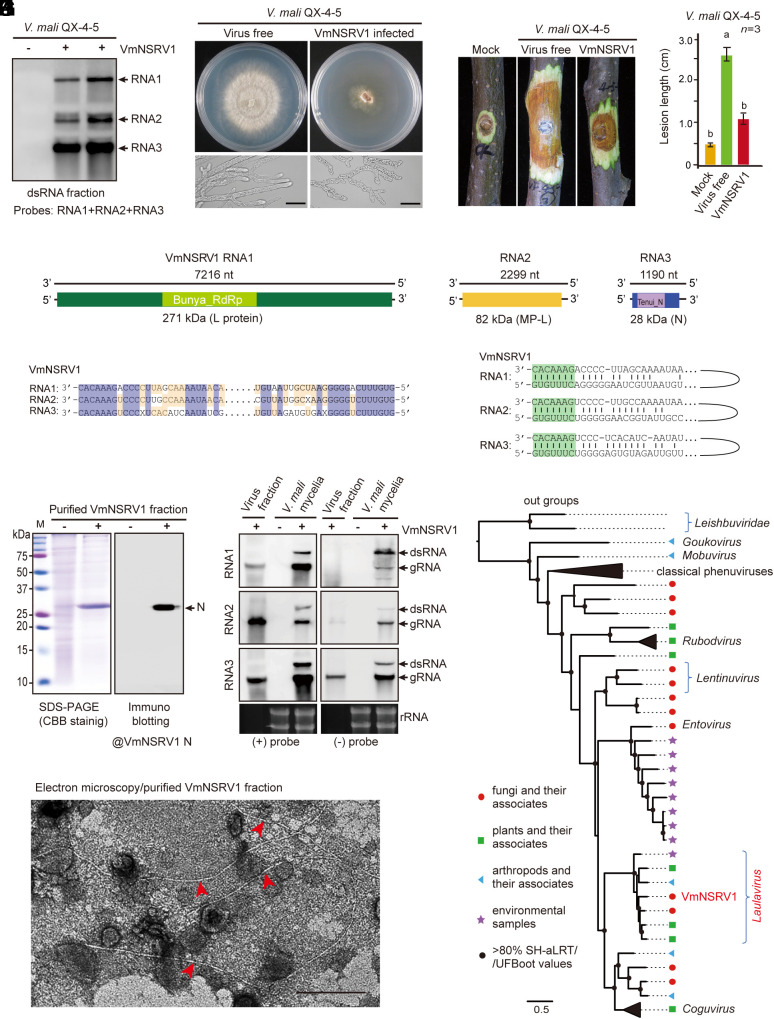
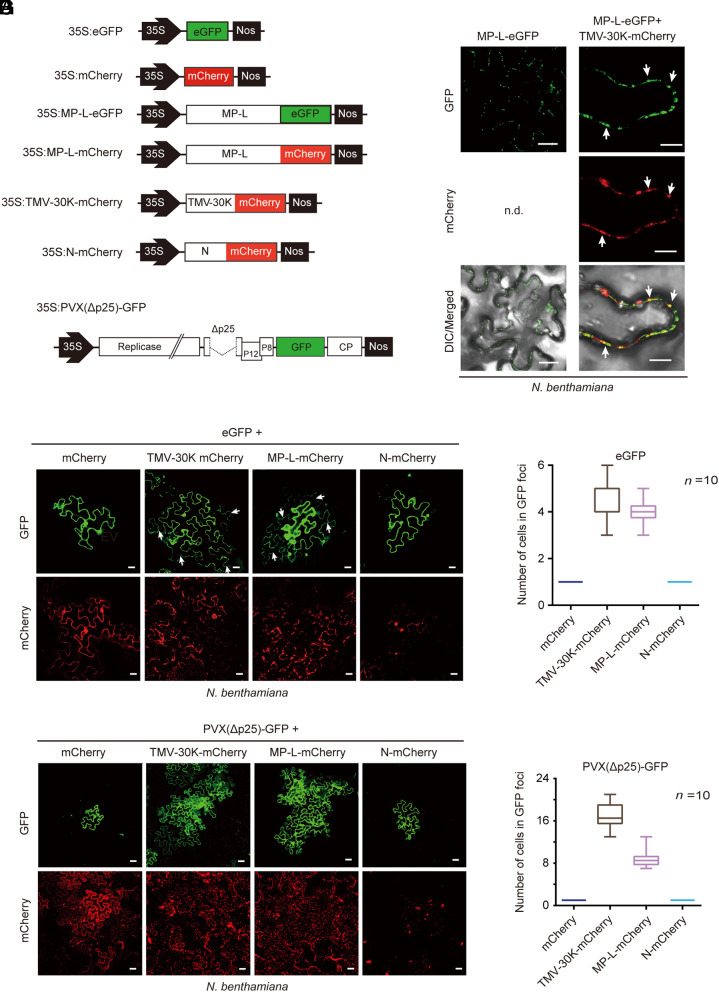
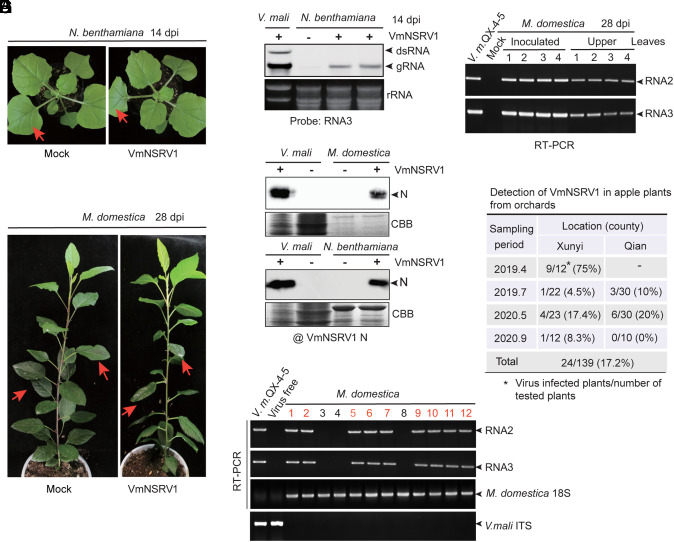
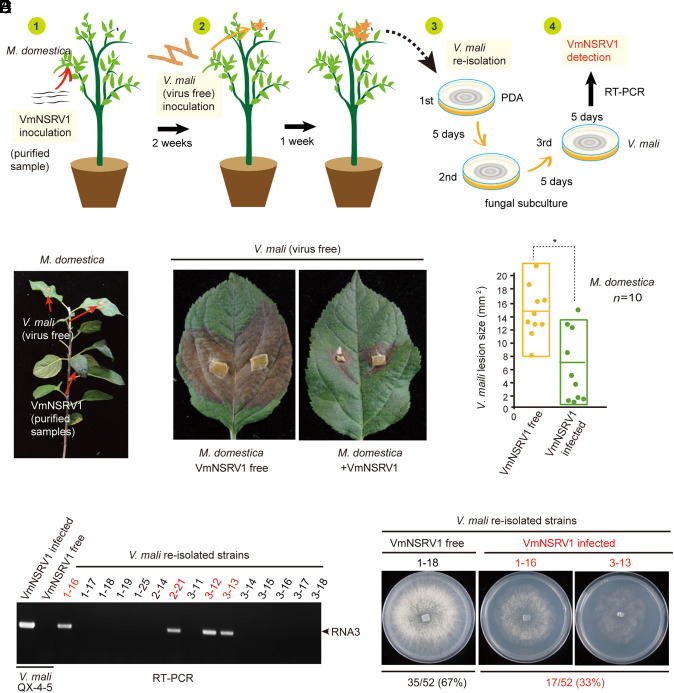
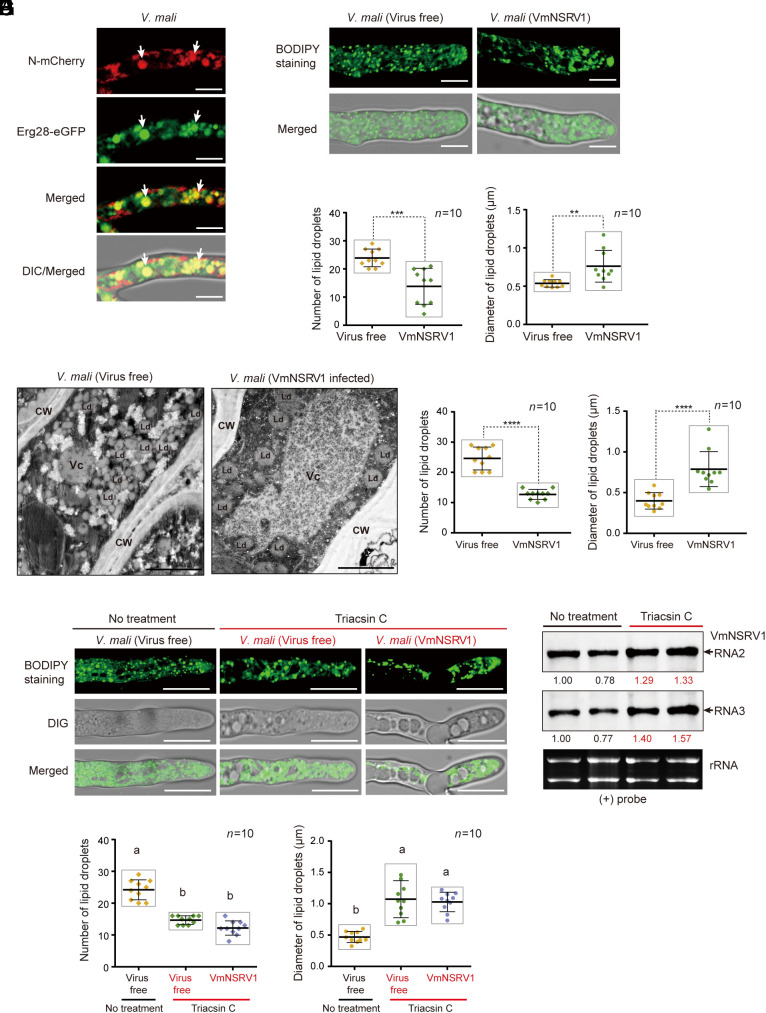
The presence of viruses that spread to both plant and fungal populations in nature has posed intriguingly scientific question. We found a negative-strand RNA virus related to members of the family Phenuiviridae, named Valsa mali negative-strand RNA virus 1 (VmNSRV1), which induced strong hypovirulence and was prevalent in a population of the phytopathogenic fungus of apple Valsa canker (Valsa mali) infecting apple orchards in the Shaanxi Province of China. Intriguingly, VmNSRV1 encodes a protein with a viral cell-to-cell movement function in plant tissue. Mechanical leaf inoculation showed that VmNSRV1 could systemically infect plants. Moreover, VmNSRV1 was detected in 24 out of 139 apple trees tested in orchards in Shaanxi Province. Fungal inoculation experiments showed that VmNSRV1 could be bidirectionally transmitted between apple plants and V. mali, and VmNSRV1 infection in plants reduced the development of fungal lesions on leaves. Additionally, the nucleocapsid protein encoded by VmNSRV1 is associated with and rearranged lipid droplets in both fungal and plant cells. VmNSRV1 represents a virus that has adapted and spread to both plant and fungal hosts and shuttles between these two organisms in nature (phyto-mycovirus) and is potential to be utilized for the biocontrol method against plant fungal diseases. This finding presents further insights into the virus evolution and adaptation encompassing both plant and fungal hosts.
Keywords: cross-kingdom infection; hypovirulence; movement protein; mycovirus; negative-strand RNA virus.
Conflict of interest statement
Competing interests statement:The authors declare no competing interest.
Figures
References
-
- Pommerville J. C., Alcamo’s Fundamentals of Microbiology: Body Systems (Jones & Bartlett Publishers, 2012).
-
- Sun L., Kondo H., Andika I. B., “Cross-kingdom virus infection” in Encyclopedia of Virology, Bamford D. H., Zuckerman M., Eds. (Elsevier, Amsterdam, 2020), pp. 443–449.
-
- Navaratnarajah C., Warrier R., Kuhn R. J., “Assembly of viruses: Enveloped particles” in Encyclopedia of Virology, van Regenmortel M. H. V., Mahy B. W. J., Eds. (Elsevier, Amsterdam, 2008), pp. 193–200, 10.1016/B978-012374410-4.00667-1. - DOI
-
- Whitfield A. E., Falk B. W., Rotenberg D., Insect vector-mediated transmission of plant viruses. Virology 479, 278–289 (2015). - PubMed
MeSH terms
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
