

NIa-Pro of sugarcane mosaic virus targets Corn Cysteine Protease 1 (CCP1) to undermine salicylic acid-mediated defense in maize
- PMID: 38484013
- PMCID: PMC10965072
- DOI: 10.1371/journal.ppat.1012086
NIa-Pro of sugarcane mosaic virus targets Corn Cysteine Protease 1 (CCP1) to undermine salicylic acid-mediated defense in maize
Abstract
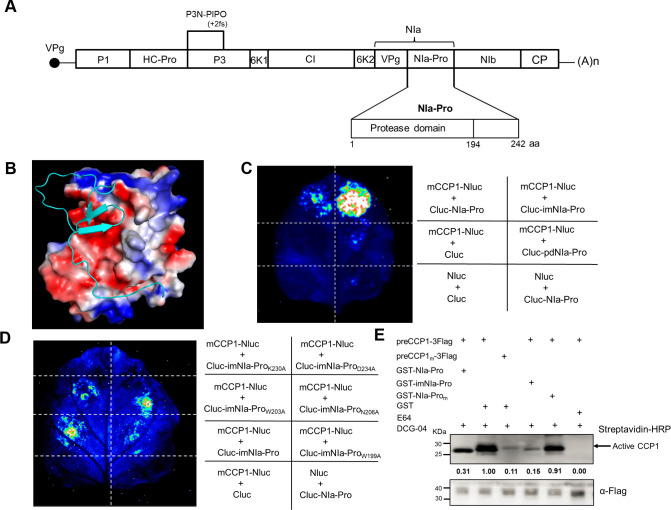
Papain-like cysteine proteases (PLCPs) play pivotal roles in plant defense against pathogen invasions. While pathogens can secrete effectors to target and inhibit PLCP activities, the roles of PLCPs in plant-virus interactions and the mechanisms through which viruses neutralize PLCP activities remain largely uncharted. Here, we demonstrate that the expression and activity of a maize PLCP CCP1 (Corn Cysteine Protease), is upregulated following sugarcane mosaic virus (SCMV) infection. Transient silencing of CCP1 led to a reduction in PLCP activities, thereby promoting SCMV infection in maize. Furthermore, the knockdown of CCP1 resulted in diminished salicylic acid (SA) levels and suppressed expression of SA-responsive pathogenesis-related genes. This suggests that CCP1 plays a role in modulating the SA signaling pathway. Interestingly, NIa-Pro, the primary protease of SCMV, was found to interact with CCP1, subsequently inhibiting its protease activity. A specific motif within NIa-Pro termed the inhibitor motif was identified as essential for its interaction with CCP1 and the suppression of its activity. We have also discovered that the key amino acids responsible for the interaction between NIa-Pro and CCP1 are crucial for the virulence of SCMV. In conclusion, our findings offer compelling evidence that SCMV undermines maize defense mechanisms through the interaction of NIa-Pro with CCP1. Together, these findings shed a new light on the mechanism(s) controlling the arms races between virus and plant.
Copyright: © 2024 Yuan et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures








Similar articles
-
Maize phenylalanine ammonia-lyases contribute to resistance to Sugarcane mosaic virus infection, most likely through positive regulation of salicylic acid accumulation.Mol Plant Pathol. 2019 Oct;20(10):1365-1378. doi: 10.1111/mpp.12817. Epub 2019 Sep 5. Mol Plant Pathol. 2019. PMID: 31487111 Free PMC article.
-
A maize triacylglycerol lipase inhibits sugarcane mosaic virus infection.Plant Physiol. 2022 Jun 1;189(2):754-771. doi: 10.1093/plphys/kiac126. Plant Physiol. 2022. PMID: 35294544 Free PMC article.
-
Characterization of Maize miRNAs in Response to Synergistic Infection of Maize Chlorotic Mottle Virus and Sugarcane Mosaic Virus.Int J Mol Sci. 2019 Jun 27;20(13):3146. doi: 10.3390/ijms20133146. Int J Mol Sci. 2019. PMID: 31252649 Free PMC article.
-
Comparative proteomic analysis of the plant-virus interaction in resistant and susceptible ecotypes of maize infected with sugarcane mosaic virus.J Proteomics. 2013 Aug 26;89:124-40. doi: 10.1016/j.jprot.2013.06.005. Epub 2013 Jun 14. J Proteomics. 2013. PMID: 23770298
-
3C-like proteases at the interface of plant-virus-vector interactions: Focus on potyvirid NIa proteases and secovirid proteases.Virology. 2025 Jan;602:110299. doi: 10.1016/j.virol.2024.110299. Epub 2024 Nov 14. Virology. 2025. PMID: 39579507 Review.
Cited by
-
Genome-wide molecular characterization and expression profiling of the cysteine protease gene family in maize.BMC Genomics. 2025 Sep 1;26(1):789. doi: 10.1186/s12864-025-12003-z. BMC Genomics. 2025. PMID: 40890597 Free PMC article.
-
Identification of maize genes that condition early systemic infection of sugarcane mosaic virus through single-cell transcriptomics.Plant Commun. 2025 May 12;6(5):101297. doi: 10.1016/j.xplc.2025.101297. Epub 2025 Mar 4. Plant Commun. 2025. PMID: 40045576 Free PMC article.
-
NbPIRIN promotes the protease activity of papain-like cysteine protease NbRD21 to inhibit Chinese wheat mosaic virus infection.PLoS Pathog. 2025 Apr 2;21(4):e1013037. doi: 10.1371/journal.ppat.1013037. eCollection 2025 Apr. PLoS Pathog. 2025. PMID: 40173196 Free PMC article.
-
Carbene-Catalyzed Phthalide Ether Functionalization for Discovering Chiral Phytovirucide that Specifically Targets Viral Nia Protein to Inhibit Proliferation.Research (Wash D C). 2025 Mar 14;8:0637. doi: 10.34133/research.0637. eCollection 2025. Research (Wash D C). 2025. PMID: 40093974 Free PMC article.
References
-
- Kaschani F, Shabab M, Bozkurt T, Shindo T, Schornack S, Gu C, et al.. An effector-targeted protease contributes to defense against Phytophthora infestans and is under diversifying selection in natural hosts. Plant Physiol. 2010;154(4):1794–804. doi: 10.1104/pp.110.158030 WOS:000284834000017. - DOI - PMC - PubMed
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
