

Emergence and maintenance of stable coexistence during a long-term multicellular evolution experiment
- PMID: 38486107
- PMCID: PMC11090753
- DOI: 10.1038/s41559-024-02367-y
Emergence and maintenance of stable coexistence during a long-term multicellular evolution experiment
Abstract
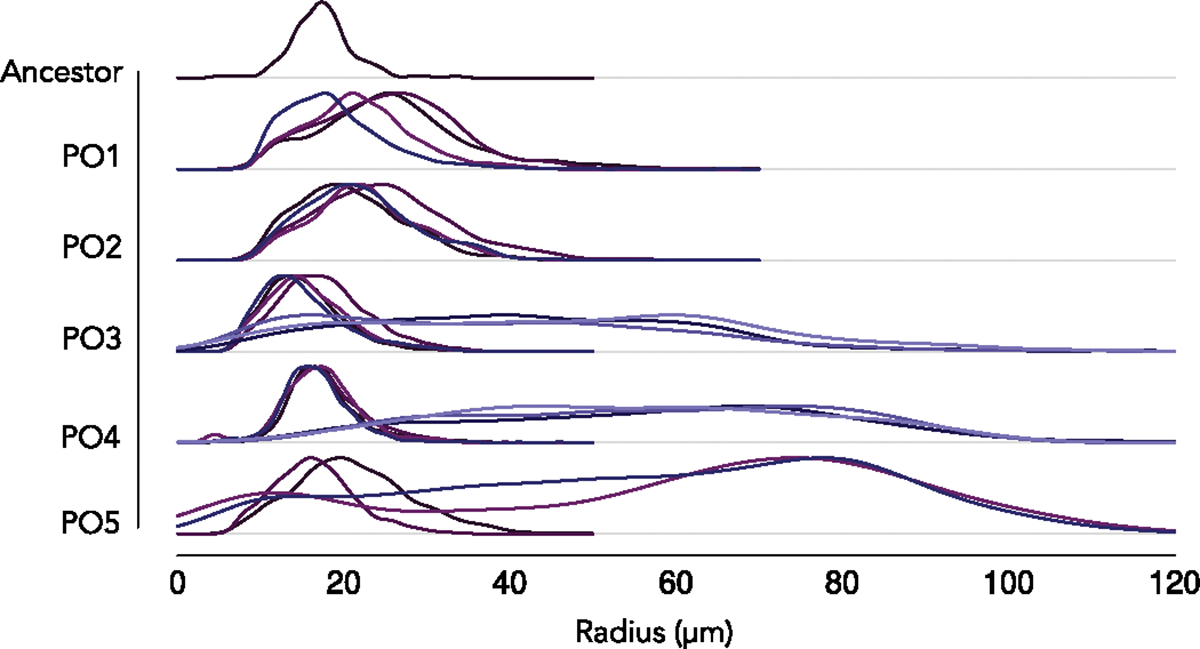
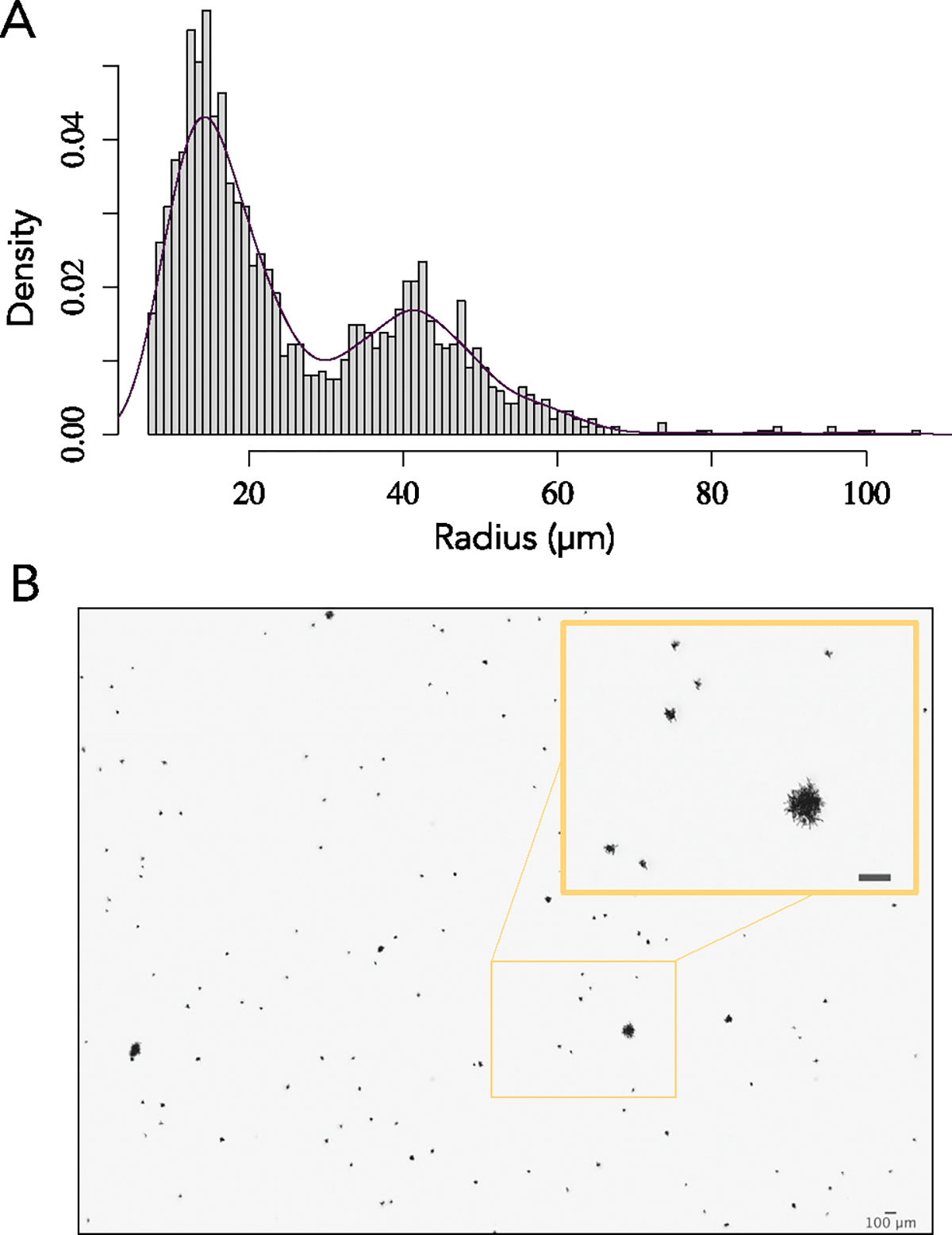
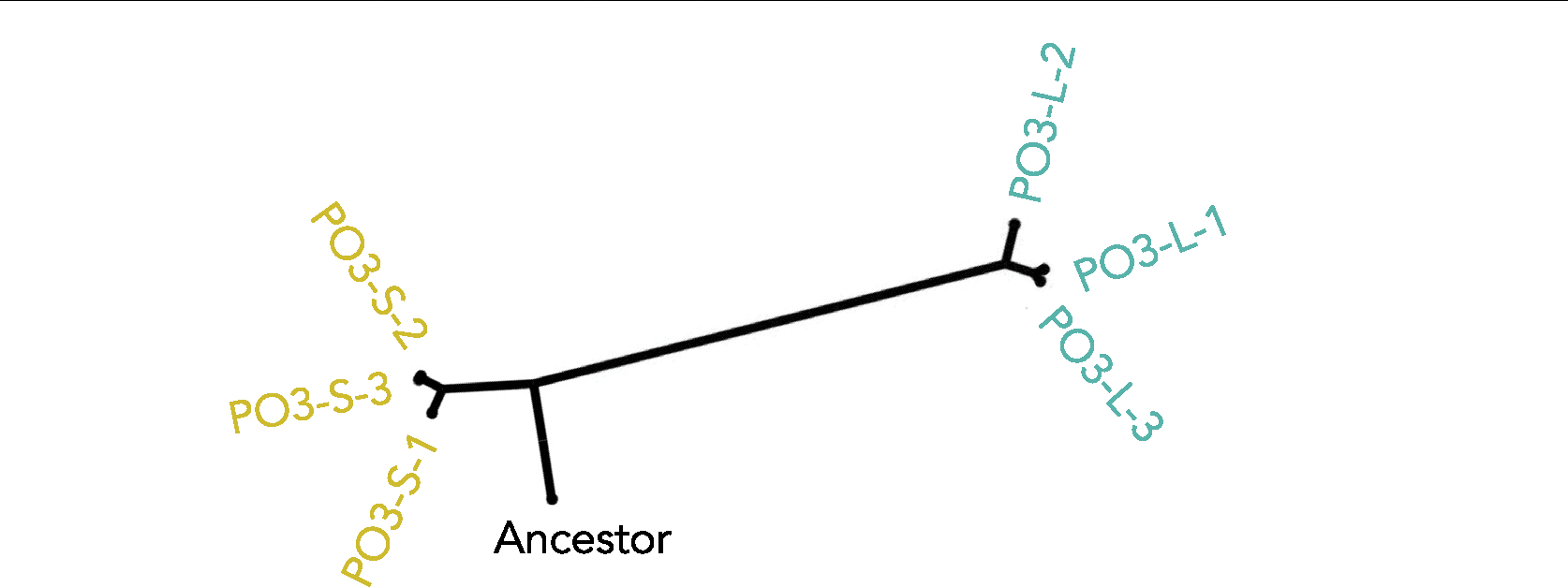
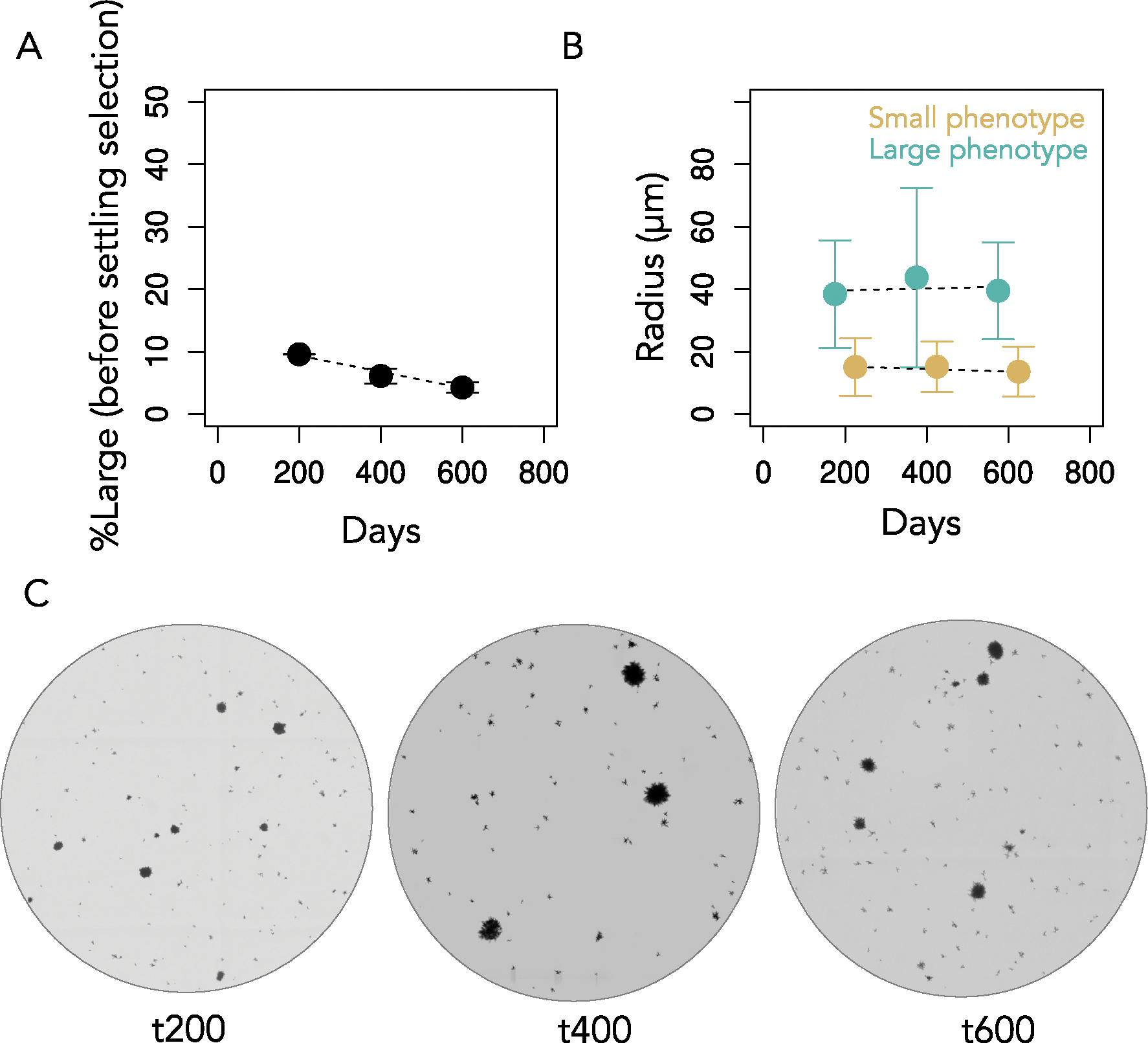
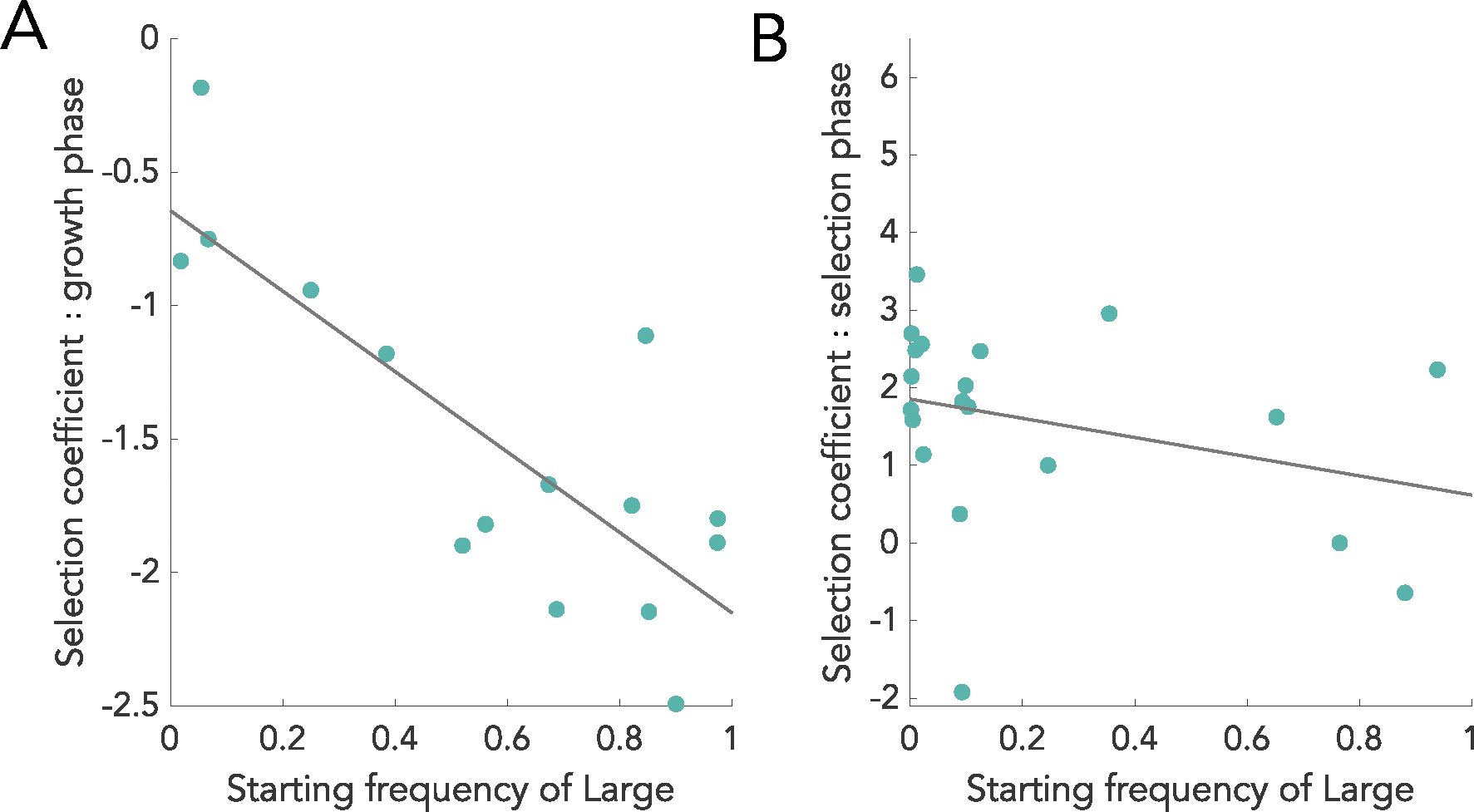
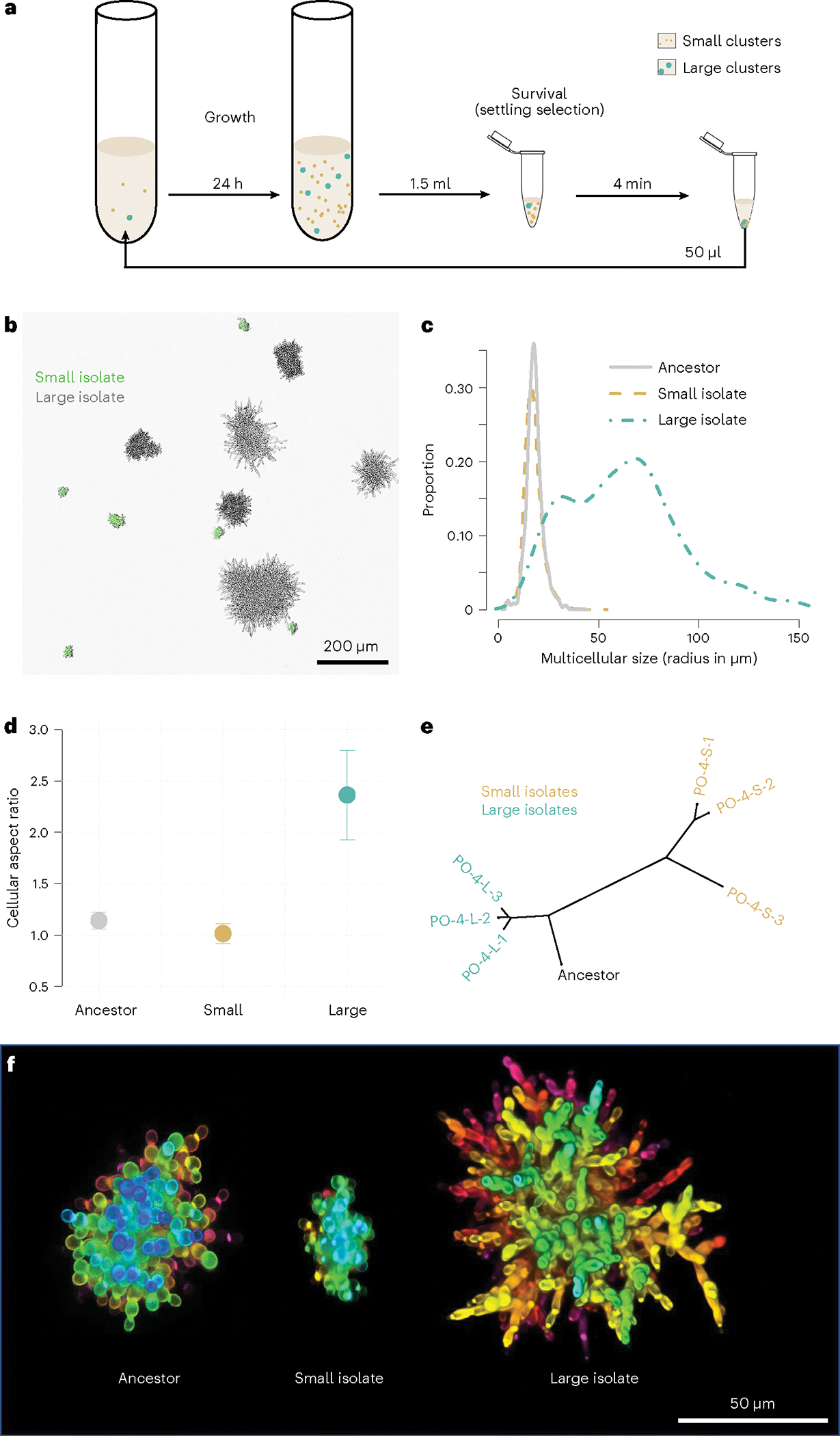
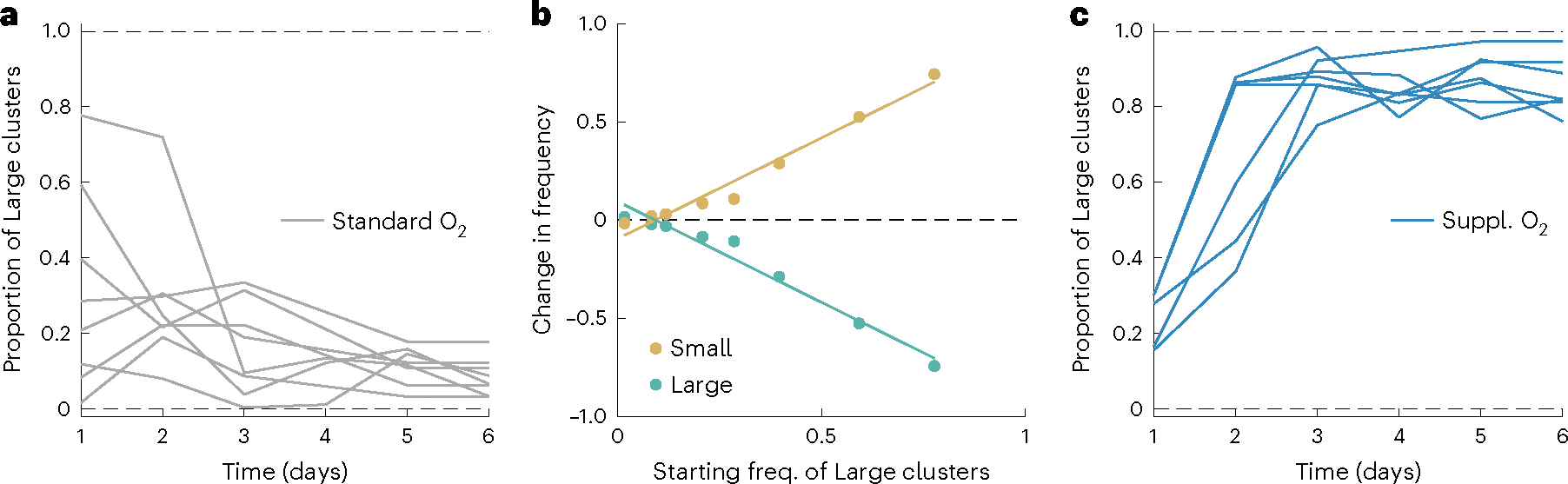
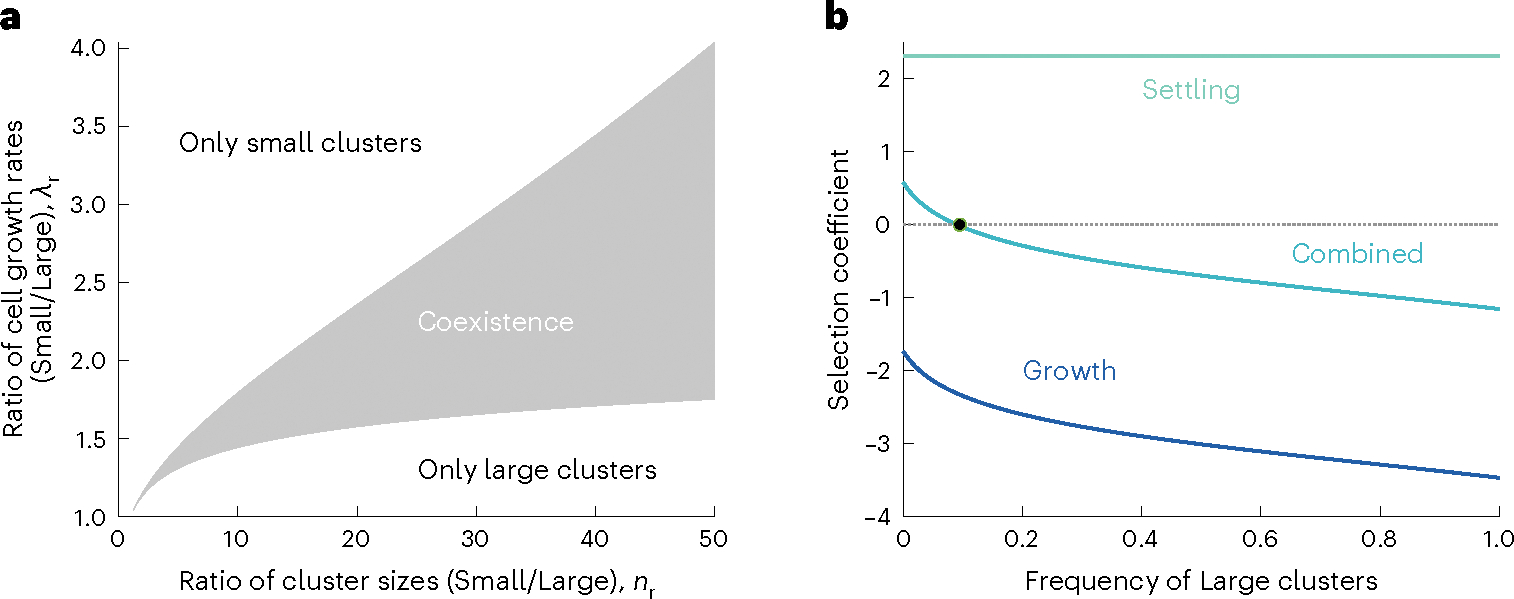
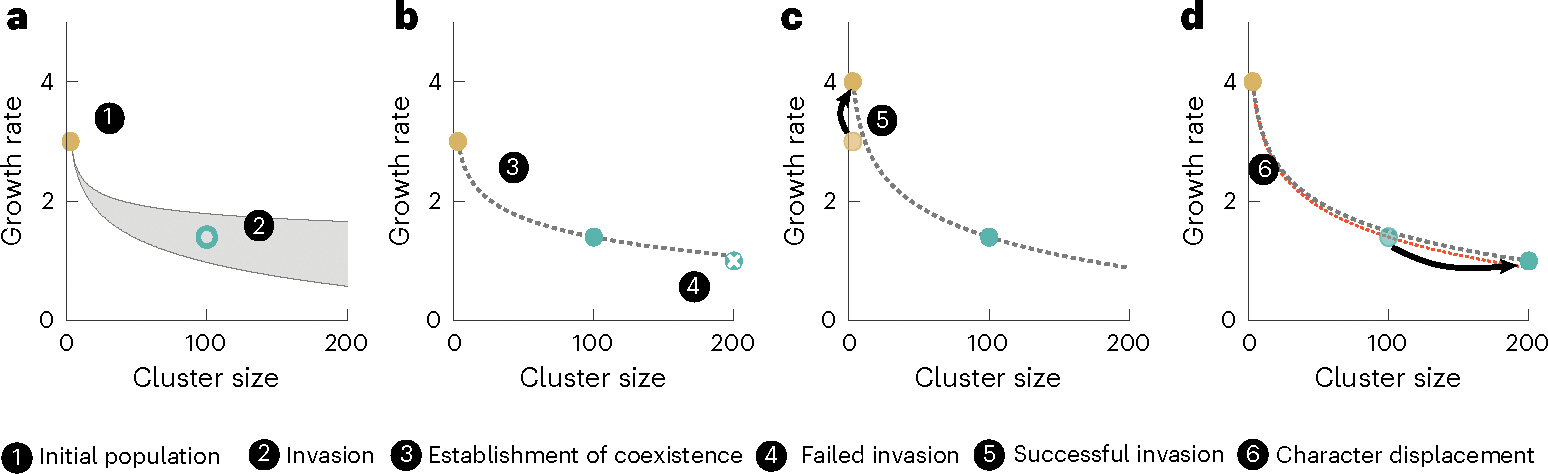
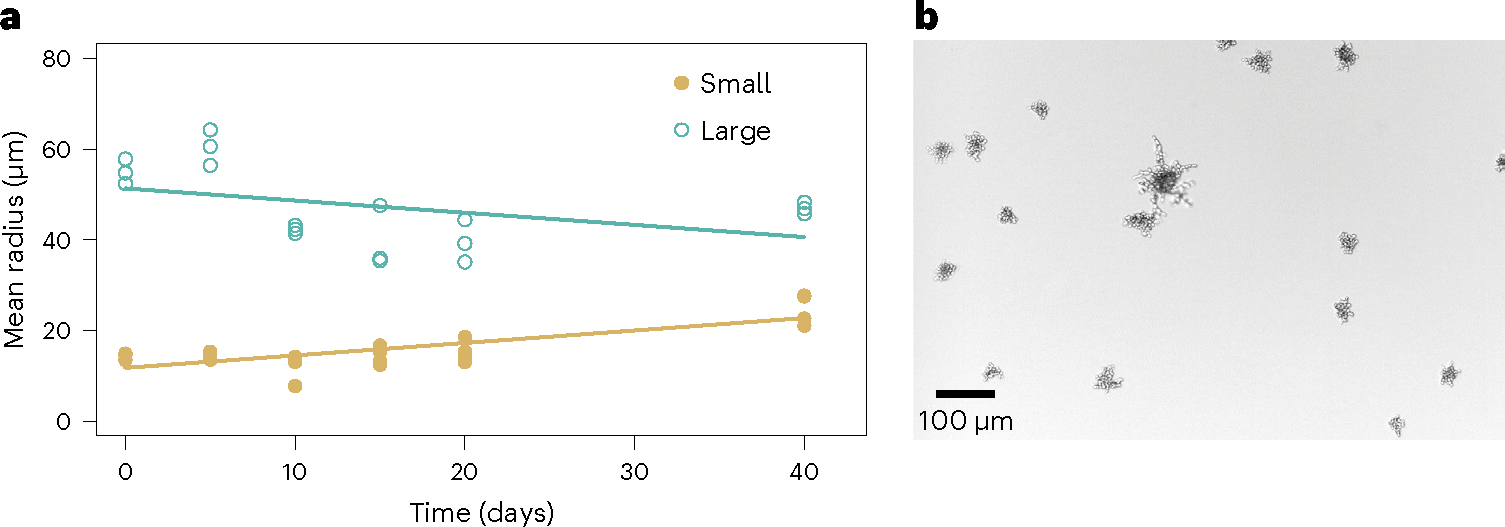
The evolution of multicellular life spurred evolutionary radiations, fundamentally changing many of Earth's ecosystems. Yet little is known about how early steps in the evolution of multicellularity affect eco-evolutionary dynamics. Through long-term experimental evolution, we observed niche partitioning and the adaptive divergence of two specialized lineages from a single multicellular ancestor. Over 715 daily transfers, snowflake yeast were subjected to selection for rapid growth, followed by selection favouring larger group size. Small and large cluster-forming lineages evolved from a monomorphic ancestor, coexisting for over ~4,300 generations, specializing on divergent aspects of a trade-off between growth rate and survival. Through modelling and experimentation, we demonstrate that coexistence is maintained by a trade-off between organismal size and competitiveness for dissolved oxygen. Taken together, this work shows how the evolution of a new level of biological individuality can rapidly drive adaptive diversification and the expansion of a nascent multicellular niche, one of the most historically impactful emergent properties of this evolutionary transition.
© 2024. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures
Update of
-
Emergence and maintenance of stable coexistence during a long-term multicellular evolution experiment.bioRxiv [Preprint]. 2023 Jan 20:2023.01.19.524803. doi: 10.1101/2023.01.19.524803. bioRxiv. 2023. Update in: Nat Ecol Evol. 2024 May;8(5):1010-1020. doi: 10.1038/s41559-024-02367-y. PMID: 36711513 Free PMC article. Updated. Preprint.
References
-
- Maloof AC et al. The earliest Cambrian record of animals and ocean geochemical change. GSA Bull. 122, 1731–1774 (2010).
-
- McMahon S & Parnell J The deep history of Earth’s biomass. J. Geol. Soc. 175, 716–720 (2018).
-
- Delwiche CF & Cooper ED The evolutionary origin of a terrestrial flora. Curr. Biol. 25, R899–R910 (2015). - PubMed
-
- Odling-Smee FJ, Laland KN & Feldman MW Niche construction. Am. Nat. 147, 641–648 (1996).
-
- Boyce CK The evolutionary history of roots and leaves. in Vascular Transport in Plants (eds Holbrook NM & Zwieniecki MA) 479–499 (Elsevier, 2005).
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
