

Identification of membrane curvature sensing motifs essential for VPS37A phagophore recruitment and autophagosome closure
- PMID: 38491121
- PMCID: PMC10942982
- DOI: 10.1038/s42003-024-06026-7
Identification of membrane curvature sensing motifs essential for VPS37A phagophore recruitment and autophagosome closure
Abstract
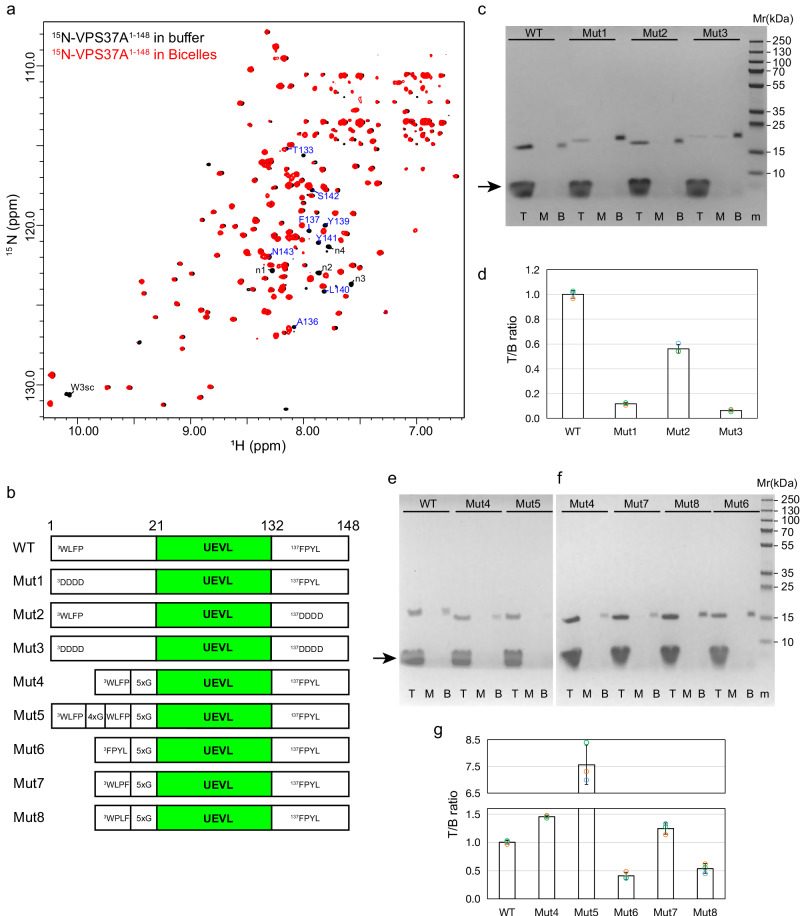
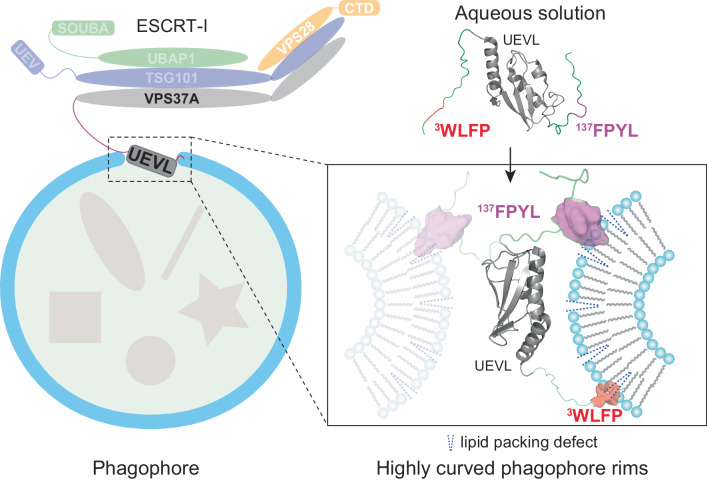
VPS37A, an ESCRT-I complex component, is required for recruiting a subset of ESCRT proteins to the phagophore for autophagosome closure. However, the mechanism by which VPS37A is targeted to the phagophore remains obscure. Here, we demonstrate that the VPS37A N-terminal domain exhibits selective interactions with highly curved membranes, mediated by two membrane-interacting motifs within the disordered regions surrounding its Ubiquitin E2 variant-like (UEVL) domain. Site-directed mutations of residues in these motifs disrupt ESCRT-I localization to the phagophore and result in defective phagophore closure and compromised autophagic flux in vivo, highlighting their essential role during autophagy. In conjunction with the UEVL domain, we postulate that these motifs guide a functional assembly of the ESCRT machinery at the highly curved tip of the phagophore for autophagosome closure. These results advance the notion that the distinctive membrane architecture of the cup-shaped phagophore spatially regulates autophagosome biogenesis.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures



Similar articles
-
Unveiling the physiological impact of ESCRT-dependent autophagosome closure by targeting the VPS37A ubiquitin E2 variant-like domain.Cell Rep. 2024 Dec 24;43(12):115016. doi: 10.1016/j.celrep.2024.115016. Epub 2024 Nov 27. Cell Rep. 2024. PMID: 39607828 Free PMC article.
-
Blocking autophagosome closure manifests the roles of mammalian Atg8-family proteins in phagophore formation and expansion during nutrient starvation.Autophagy. 2025 May;21(5):1059-1074. doi: 10.1080/15548627.2024.2443300. Epub 2025 Jan 20. Autophagy. 2025. PMID: 39694607 Free PMC article.
-
VPS37A directs ESCRT recruitment for phagophore closure.J Cell Biol. 2019 Oct 7;218(10):3336-3354. doi: 10.1083/jcb.201902170. Epub 2019 Sep 13. J Cell Biol. 2019. PMID: 31519728 Free PMC article.
-
ESCRT and autophagies: Endosomal functions and beyond.Semin Cell Dev Biol. 2018 Feb;74:21-28. doi: 10.1016/j.semcdb.2017.08.014. Epub 2017 Aug 12. Semin Cell Dev Biol. 2018. PMID: 28807884 Review.
-
Lipids and membrane-associated proteins in autophagy.Protein Cell. 2021 Jul;12(7):520-544. doi: 10.1007/s13238-020-00793-9. Epub 2020 Nov 5. Protein Cell. 2021. PMID: 33151516 Free PMC article. Review.
Cited by
-
Unveiling the physiological impact of ESCRT-dependent autophagosome closure by targeting the VPS37A ubiquitin E2 variant-like domain.Cell Rep. 2024 Dec 24;43(12):115016. doi: 10.1016/j.celrep.2024.115016. Epub 2024 Nov 27. Cell Rep. 2024. PMID: 39607828 Free PMC article.
-
VPS37A Activates the Autophagy-Lysosomal Pathway for TNFR1 Degradation and Induces NF-κB-Regulated Cell Death under Metabolic Stress in Colorectal Cancer.Oncol Res. 2025 Jul 18;33(8):2085-2105. doi: 10.32604/or.2025.065739. eCollection 2025. Oncol Res. 2025. PMID: 40746890 Free PMC article.
-
The green ESCRTs: Newly defined roles for ESCRT proteins in plants.J Biol Chem. 2025 May;301(5):108465. doi: 10.1016/j.jbc.2025.108465. Epub 2025 Mar 27. J Biol Chem. 2025. PMID: 40157538 Free PMC article. Review.
-
The expanding repertoire of ESCRT functions in cell biology and disease.Nature. 2025 Jun;642(8069):877-888. doi: 10.1038/s41586-025-08950-y. Epub 2025 Jun 25. Nature. 2025. PMID: 40562928 Review.
-
Blocking autophagosome closure manifests the roles of mammalian Atg8-family proteins in phagophore formation and expansion during nutrient starvation.Autophagy. 2025 May;21(5):1059-1074. doi: 10.1080/15548627.2024.2443300. Epub 2025 Jan 20. Autophagy. 2025. PMID: 39694607 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
