

Loss of GIPR in LEPR cells impairs glucose control by GIP and GIP:GLP-1 co-agonism without affecting body weight and food intake in mice
- PMID: 38492844
- PMCID: PMC10973979
- DOI: 10.1016/j.molmet.2024.101915
Loss of GIPR in LEPR cells impairs glucose control by GIP and GIP:GLP-1 co-agonism without affecting body weight and food intake in mice
Abstract
Objective: The glucose-dependent insulinotropic polypeptide (GIP) decreases body weight via central GIP receptor (GIPR) signaling, but the underlying mechanisms remain largely unknown. Here, we assessed whether GIP regulates body weight and glucose control via GIPR signaling in cells that express the leptin receptor (Lepr).
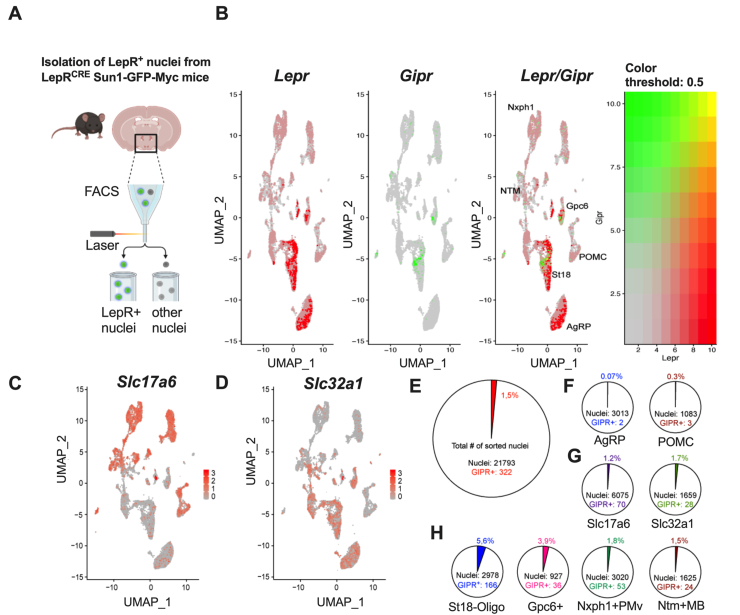
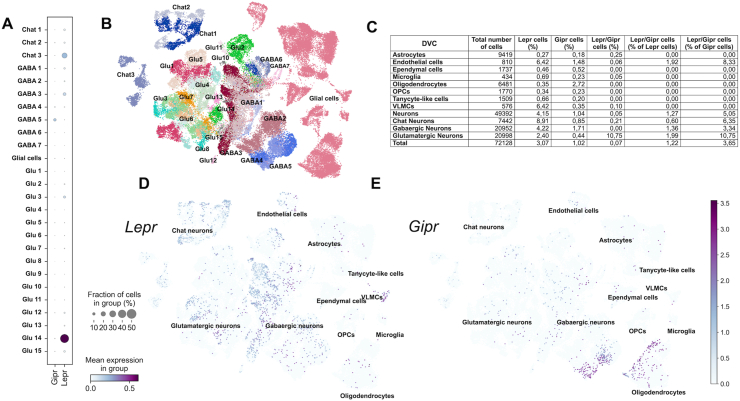
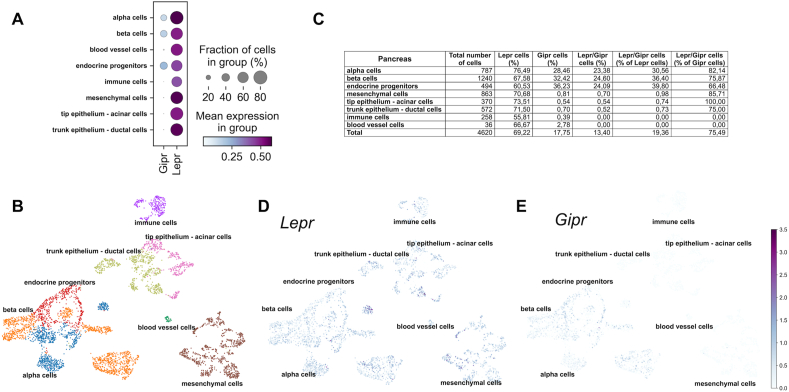
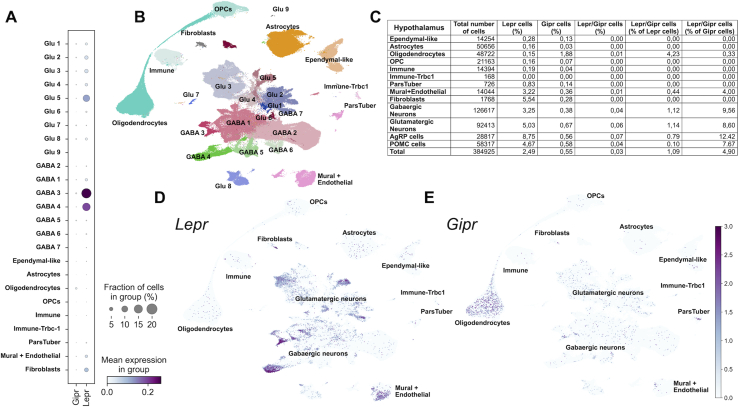
Methods: Hypothalamic, hindbrain, and pancreatic co-expression of Gipr and Lepr was assessed using single cell RNAseq analysis. Mice with deletion of Gipr in Lepr cells were generated and metabolically characterized for alterations in diet-induced obesity (DIO), glucose control and leptin sensitivity. Long-acting single- and dual-agonists at GIPR and GLP-1R were further used to assess drug effects on energy and glucose metabolism in DIO wildtype (WT) and Lepr-Gipr knock-out (KO) mice.
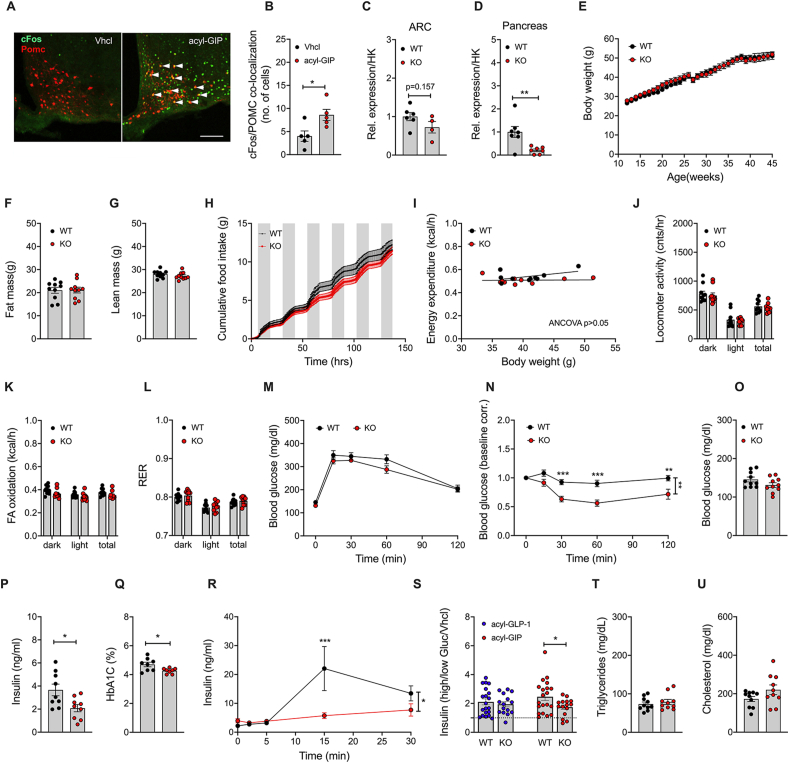
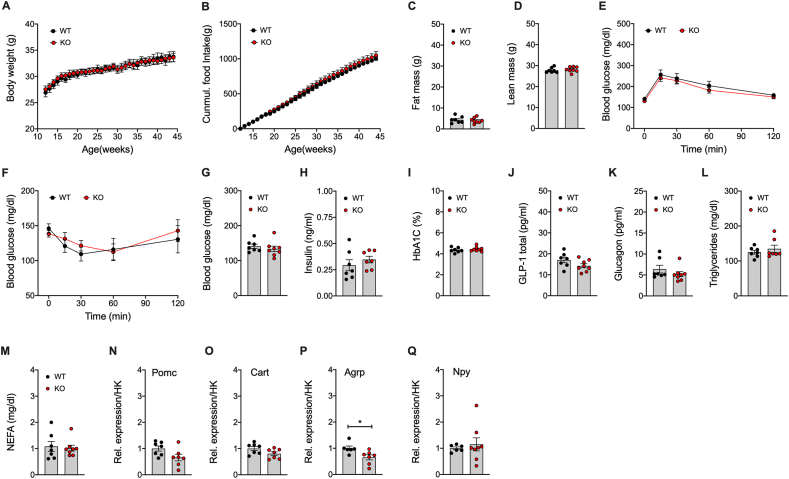
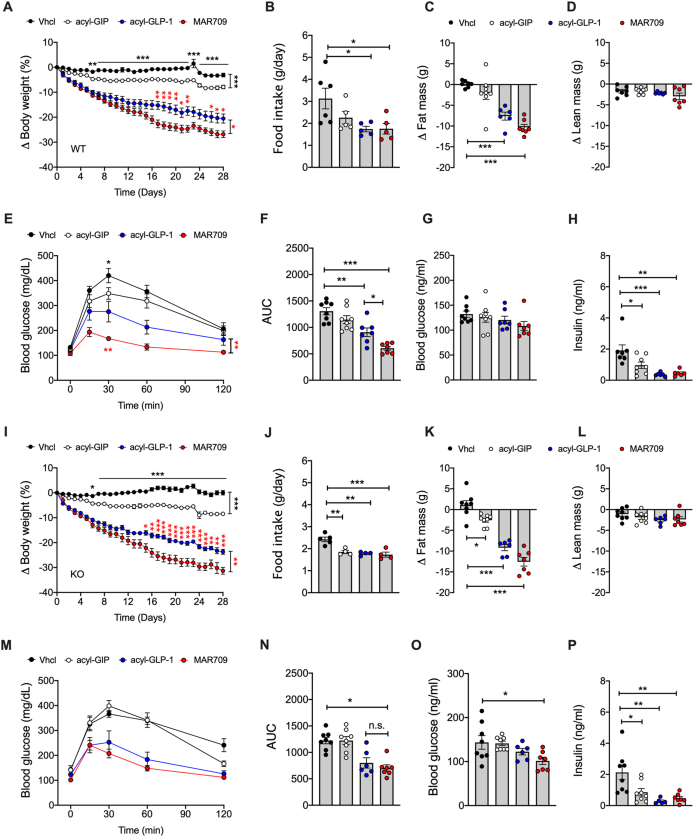
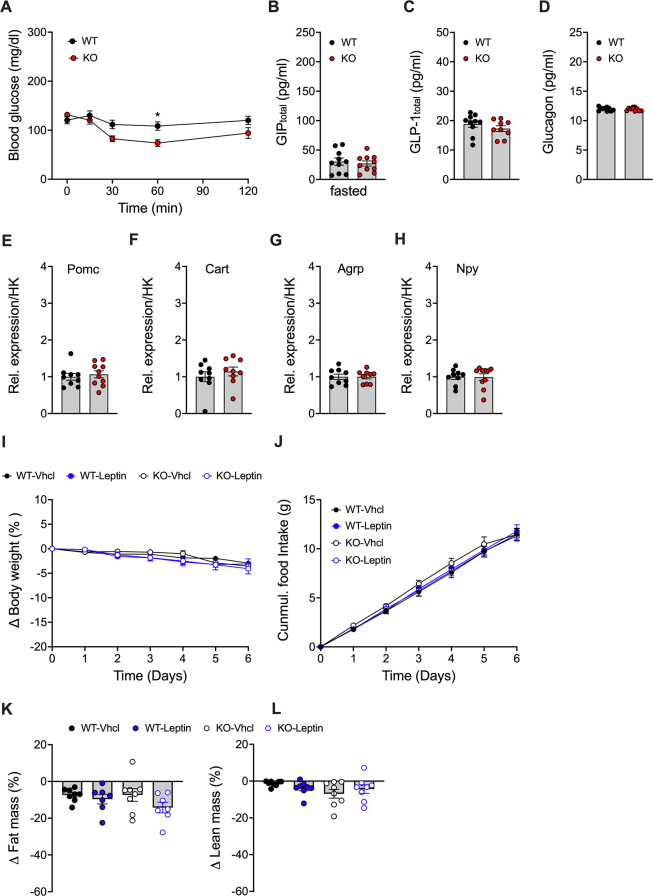
Results: Gipr and Lepr show strong co-expression in the pancreas, but not in the hypothalamus and hindbrain. DIO Lepr-Gipr KO mice are indistinguishable from WT controls related to body weight, food intake and diet-induced leptin resistance. Acyl-GIP and the GIPR:GLP-1R co-agonist MAR709 remain fully efficacious to decrease body weight and food intake in DIO Lepr-Gipr KO mice. Consistent with the demonstration that Gipr and Lepr highly co-localize in the endocrine pancreas, including the β-cells, we find the superior glycemic effect of GIPR:GLP-1R co-agonism over single GLP-1R agonism to vanish in Lepr-Gipr KO mice.
Conclusions: GIPR signaling in cells/neurons that express the leptin receptor is not implicated in the control of body weight or food intake, but is of crucial importance for the superior glycemic effects of GIPR:GLP-1R co-agonism relative to single GLP-1R agonism.
Keywords: GIP; GIPR:GLP-1R co-agonism; GLP-1; Obesity; Type 2 diabetes.
Copyright © 2024 The Author(s). Published by Elsevier GmbH.. All rights reserved.
Conflict of interest statement
Declaration of competing interest MHT is a member of the scientific advisory board of ERX Pharmaceuticals, Cambridge, Mass. He was a member of the Research Cluster Advisory Panel (ReCAP) of the Novo Nordisk Foundation between 2017 and 2019. He attended a scientific advisory board meeting of the Novo Nordisk Foundation Center for Basic Metabolic Research, University of Copenhagen, in 2016. He received funding for his research projects by Novo Nordisk (2016–2020) and Sanofi-Aventis (2012–2019). He was a consultant for Bionorica SE (2013–2017), Menarini Ricerche S.p.A. (2016), and Bayer Pharma AG Berlin (2016). As former Director of the Helmholtz Diabetes Center and the Institute for Diabetes and Obesity at Helmholtz Zentrum München (2011–2018), and since 2018, as CEO of Helmholtz Zentrum München, he has been responsible for collaborations with a multitude of companies and institutions, worldwide. In this capacity, he discussed potential projects with and has signed/signs contracts for his institute(s) and for the staff for research funding and/or collaborations with industry and academia, worldwide, including but not limited to pharmaceutical corporations like Boehringer Ingelheim, Eli Lilly, Novo Nordisk, Medigene, Arbormed, BioSyngen, and others. In this role, he was/is further responsible for commercial technology transfer activities of his institute(s), including diabetes related patent portfolios of Helmholtz Zentrum München as, e.g., WO/2016/188932 A2 or WO/2017/194499 A1. MHT confirms that to the best of his knowledge none of the above funding sources were involved in the preparation of this paper. TDM and K.S. receive research funding by Novo Nordisk but these funds are unrelated the here described work. DJD has received speaking and consulting fees from Merck and Novo Nordisk Inc and consulting fees from Forkhead Biopharmaceuticals and Kallyope Inc. R.D.D is a co-inventor on intellectual property owned by Indiana University and licensed to Novo Nordisk. He was previously employed by Novo Nordisk. P.J.K, S.M., and B.F. are current employees of Novo Nordisk. TDM receives funding from Novo Nordisk and received speaking fees within the last 3 years from Novo Nordisk, Eli Lilly, AstraZeneca, Merck, Berlin Chemie AG, and Mercodia.
Figures

Similar articles
-
Optimized GIP analogs promote body weight lowering in mice through GIPR agonism not antagonism.Mol Metab. 2019 Feb;20:51-62. doi: 10.1016/j.molmet.2018.12.001. Epub 2018 Dec 5. Mol Metab. 2019. PMID: 30578168 Free PMC article.
-
Glucose-dependent insulinotropic polypeptide regulates body weight and food intake via GABAergic neurons in mice.Nat Metab. 2023 Dec;5(12):2075-2085. doi: 10.1038/s42255-023-00931-7. Epub 2023 Nov 9. Nat Metab. 2023. PMID: 37946085 Free PMC article.
-
Specific loss of GIPR signaling in GABAergic neurons enhances GLP-1R agonist-induced body weight loss.Mol Metab. 2025 May;95:102074. doi: 10.1016/j.molmet.2024.102074. Epub 2024 Nov 26. Mol Metab. 2025. PMID: 39612941 Free PMC article.
-
Targeting the GIPR for obesity: To agonize or antagonize? Potential mechanisms.Mol Metab. 2021 Apr;46:101139. doi: 10.1016/j.molmet.2020.101139. Epub 2020 Dec 5. Mol Metab. 2021. PMID: 33290902 Free PMC article. Review.
-
Regulation of energy metabolism through central GIPR signaling.Peptides. 2024 Jun;176:171198. doi: 10.1016/j.peptides.2024.171198. Epub 2024 Mar 26. Peptides. 2024. PMID: 38527521 Review.
Cited by
-
A Contemporary Rationale for Agonism of the GIP Receptor in the Treatment of Obesity.Diabetes. 2025 Aug 1;74(8):1326-1333. doi: 10.2337/dbi24-0026. Diabetes. 2025. PMID: 40521890 Free PMC article. Review.
-
Glucose-dependent insulinotropic polypeptide (GIP).Mol Metab. 2025 May;95:102118. doi: 10.1016/j.molmet.2025.102118. Epub 2025 Feb 28. Mol Metab. 2025. PMID: 40024571 Free PMC article. Review.
-
Therapeutic Targeting of the GIP Receptor-Revisiting the Controversies.Diabetes. 2025 Aug 1;74(8):1320-1325. doi: 10.2337/db25-0393. Diabetes. 2025. PMID: 40521880 Free PMC article. Review.
-
GIPR agonism and antagonism decrease body weight and food intake via different mechanisms in male mice.Nat Metab. 2025 Jun;7(6):1282-1298. doi: 10.1038/s42255-025-01294-x. Epub 2025 Apr 29. Nat Metab. 2025. PMID: 40301583 Free PMC article.
References
-
- Finan B., Ma T., Ottaway N., Muller T.D., Habegger K.M., Heppner K.M., et al. Unimolecular dual incretins maximize metabolic benefits in rodents, monkeys, and humans. Sci Transl Med. 2013;5 - PubMed
-
- Heise T., Mari A., DeVries J.H., Urva S., Li J., Pratt E.J., et al. Effects of subcutaneous tirzepatide versus placebo or semaglutide on pancreatic islet function and insulin sensitivity in adults with type 2 diabetes: a multicentre, randomised, double-blind, parallel-arm, phase 1 clinical trial. Lancet Diabetes Endocrinol. 2022;10:418–429. - PubMed
-
- Frias J.P., Davies M.J., Rosenstock J., Perez Manghi F.C., Fernandez Lando L., Bergman B.K., et al. Tirzepatide versus semaglutide once weekly in patients with type 2 diabetes. N Engl J Med. 2021;385:503–515. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
