

Transcriptome regulation by PARP13 in basal and antiviral states in human cells
- PMID: 38495826
- PMCID: PMC10943485
- DOI: 10.1016/j.isci.2024.109251
Transcriptome regulation by PARP13 in basal and antiviral states in human cells
Abstract
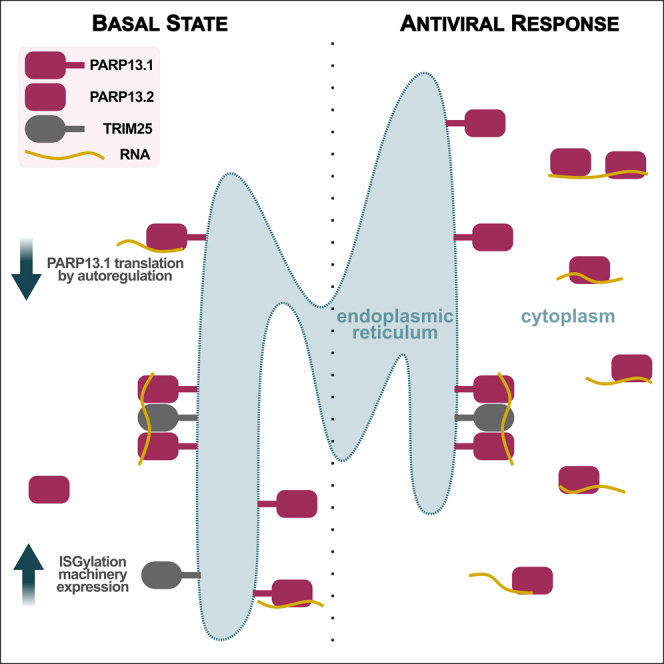
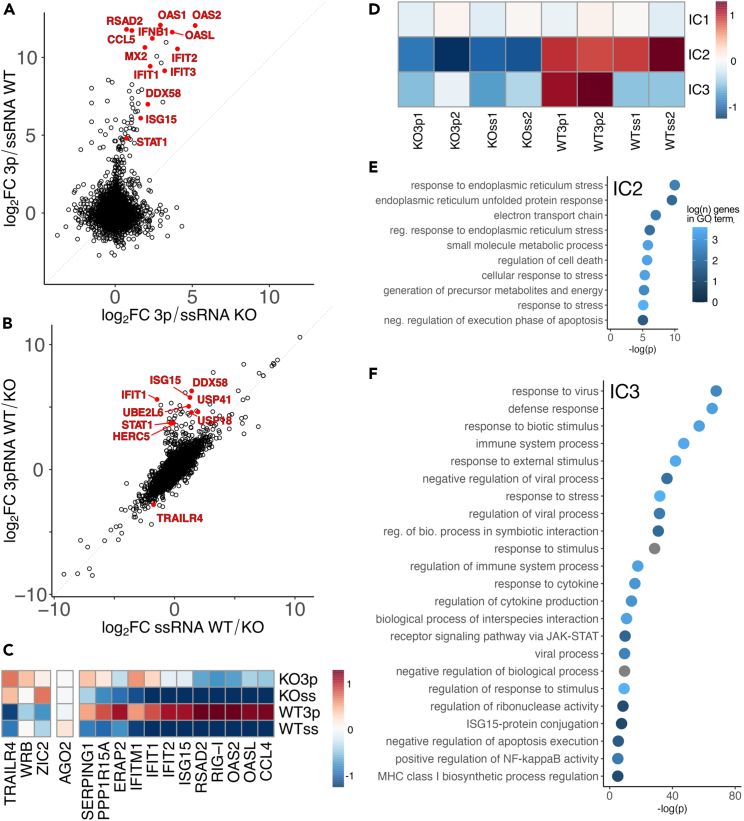
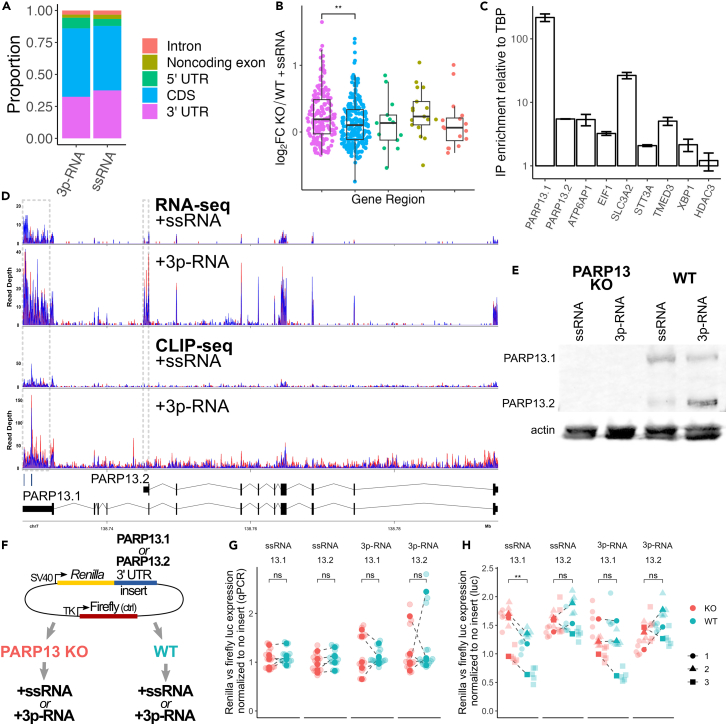
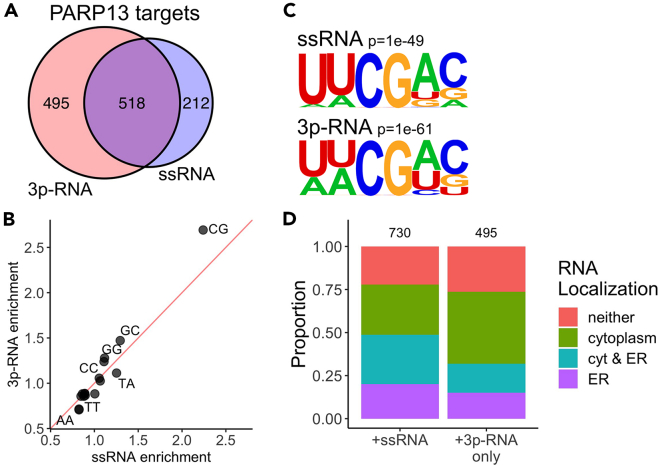
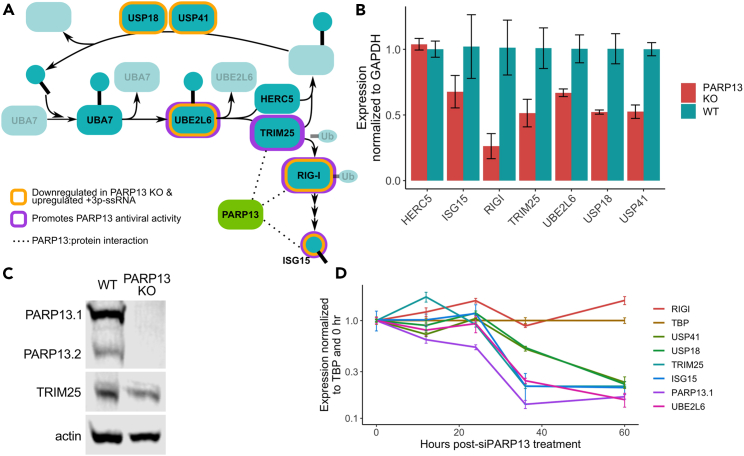
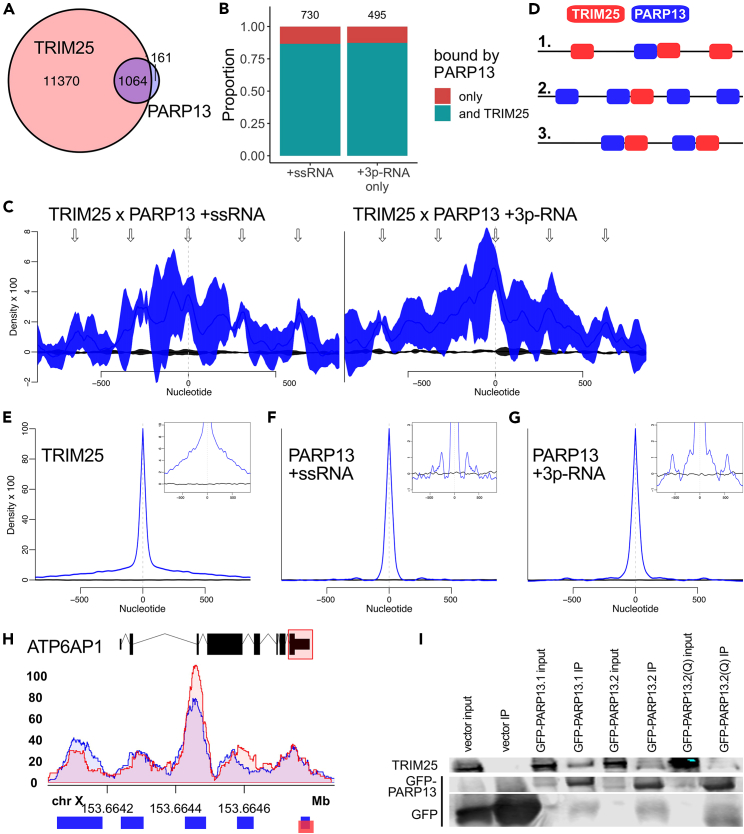
The RNA-binding protein PARP13 is a primary factor in the innate antiviral response, which suppresses translation and drives decay of bound viral and host RNA. PARP13 interacts with many proteins encoded by interferon-stimulated genes (ISG) to activate antiviral pathways including co-translational addition of ISG15, or ISGylation. We performed enhanced crosslinking immunoprecipitation (eCLIP) and RNA-seq in human cells to investigate PARP13's role in transcriptome regulation for both basal and antiviral states. We find that the antiviral response shifts PARP13 target localization, but not its binding preferences, and that PARP13 supports the expression of ISGylation-related genes, including PARP13's cofactor, TRIM25. PARP13 associates with TRIM25 via RNA-protein interactions, and we elucidate a transcriptome-wide periodicity of PARP13 binding around TRIM25. Taken together, our study implicates PARP13 in creating and maintaining a cellular environment poised for an antiviral response through limiting PARP13 translation, regulating access to distinct mRNA pools, and elevating ISGylation machinery expression.
Keywords: Cell biology; Immune response; Molecular biology.
© 2024 The Authors.
Conflict of interest statement
G.W.Y. is co-founder, member of the Board of Directors, on the SAB, equity holder, and paid consultant for Locanabio and Eclipse BioInnovations. G.W.Y. is a visiting professor at the National University of Singapore. G.W.Y.’s interest(s) have been reviewed and approved by the University of California San Diego in accordance with its conflict-of-interest policies. The authors declare no other competing interests.
Figures
Similar articles
-
UL26 Attenuates IKKβ-Mediated Induction of Interferon-Stimulated Gene (ISG) Expression and Enhanced Protein ISGylation during Human Cytomegalovirus Infection.J Virol. 2019 Nov 13;93(23):e01052-19. doi: 10.1128/JVI.01052-19. Print 2019 Dec 1. J Virol. 2019. PMID: 31534044 Free PMC article.
-
RIG-I regulates myeloid differentiation by promoting TRIM25-mediated ISGylation.Proc Natl Acad Sci U S A. 2020 Jun 23;117(25):14395-14404. doi: 10.1073/pnas.1918596117. Epub 2020 Jun 8. Proc Natl Acad Sci U S A. 2020. PMID: 32513696 Free PMC article.
-
PARP13 regulates cellular mRNA post-transcriptionally and functions as a pro-apoptotic factor by destabilizing TRAILR4 transcript.Nat Commun. 2014 Nov 10;5:5362. doi: 10.1038/ncomms6362. Nat Commun. 2014. PMID: 25382312 Free PMC article.
-
ISGylation in Innate Antiviral Immunity and Pathogen Defense Responses: A Review.Front Cell Dev Biol. 2021 Nov 25;9:788410. doi: 10.3389/fcell.2021.788410. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 34901029 Free PMC article. Review.
-
Poly(ADP-ribose) polymerase-13 and RNA regulation in immunity and cancer.Trends Mol Med. 2015 Jun;21(6):373-84. doi: 10.1016/j.molmed.2015.03.002. Epub 2015 Apr 4. Trends Mol Med. 2015. PMID: 25851173 Free PMC article. Review.
Cited by
-
Alphavirus Evasion of Zinc Finger Antiviral Protein (ZAP) Correlates with CpG Suppression in a Specific Viral nsP2 Gene Sequence.Viruses. 2023 Mar 24;15(4):830. doi: 10.3390/v15040830. Viruses. 2023. PMID: 37112813 Free PMC article.
-
Regulation of stress granule maturation and dynamics by poly(ADP-ribose) interaction with PARP13.Nat Commun. 2025 Jan 13;16(1):621. doi: 10.1038/s41467-024-55666-0. Nat Commun. 2025. PMID: 39805863 Free PMC article.
-
PARP9-PARP13-PARP14 axis tunes colorectal cancer response to radiotherapy.J Exp Clin Cancer Res. 2025 Jul 11;44(1):199. doi: 10.1186/s13046-025-03439-y. J Exp Clin Cancer Res. 2025. PMID: 40646573 Free PMC article.
-
Endogenous ZAP is associated with altered Zika virus infection phenotype.Virol J. 2024 Nov 9;21(1):285. doi: 10.1186/s12985-024-02557-x. Virol J. 2024. PMID: 39522048 Free PMC article.
References
-
- Schwerk J., Soveg F.W., Ryan A.P., Thomas K.R., Hatfield L.D., Ozarkar S., Forero A., Kell A.M., Roby J.A., So L., et al. RNA-binding protein isoforms ZAP-S and ZAP-L have distinct antiviral and immune resolution functions. Nat. Immunol. 2019;20:1610–1620. doi: 10.1038/s41590-019-0527-6. - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
