

This is a preprint.
A Panoramic View of Cell Population Dynamics in Mammalian Aging
- PMID: 38496474
- PMCID: PMC10942312
- DOI: 10.1101/2024.03.01.583001
A Panoramic View of Cell Population Dynamics in Mammalian Aging
Update in
-
A panoramic view of cell population dynamics in mammalian aging.Science. 2025 Jan 17;387(6731):eadn3949. doi: 10.1126/science.adn3949. Epub 2025 Jan 17. Science. 2025. PMID: 39607904 Free PMC article.
Abstract
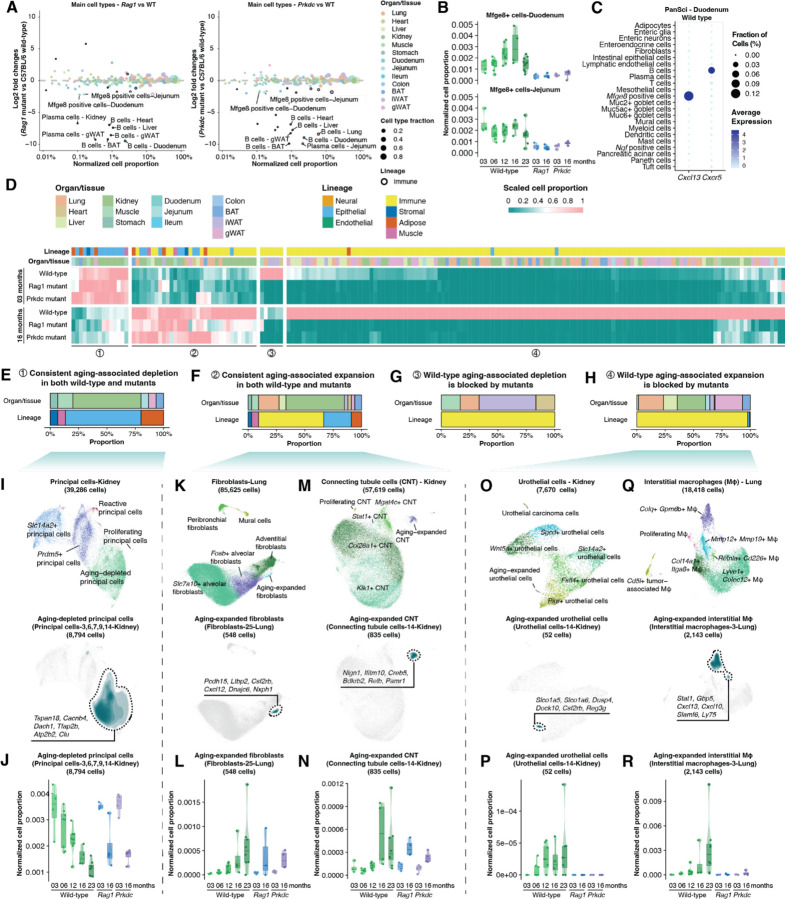
To elucidate the aging-associated cellular population dynamics throughout the body, here we present PanSci, a single-cell transcriptome atlas profiling over 20 million cells from 623 mouse tissue samples, encompassing a range of organs across different life stages, sexes, and genotypes. This comprehensive dataset allowed us to identify more than 3,000 unique cellular states and catalog over 200 distinct aging-associated cell populations experiencing significant depletion or expansion. Our panoramic analysis uncovered temporally structured, organ- and lineage-specific shifts of cellular dynamics during lifespan progression. Moreover, we investigated aging-associated alterations in immune cell populations, revealing both widespread shifts and organ-specific changes. We further explored the regulatory roles of the immune system on aging and pinpointed specific age-related cell population expansions that are lymphocyte-dependent. The breadth and depth of our 'cell-omics' methodology not only enhance our comprehension of cellular aging but also lay the groundwork for exploring the complex regulatory networks among varied cell types in the context of aging and aging-associated diseases.
Conflict of interest statement
Competing interests: In the past 3 years, R.S. has received compensation from Bristol-Myers Squibb, ImmunAI, Resolve Biosciences, Nanostring, 10x Genomics, Neptune Bio, and the NYC Pandemic Response Lab. R.S. is a co-founder and equity holder of Neptune Bio.
Figures




References
-
- Davie K., Janssens J., Koldere D., De Waegeneer M., Pech U., Kreft Ł., Aibar S., Makhzami S., Christiaens V., Bravo González-Blas C., Poovathingal S., Hulselmans G., Spanier K. I., Moerman T., Vanspauwen B., Geurs S., Voet T., Lammertyn J., Thienpont B., Liu S., Konstantinides N., Fiers M., Verstreken P., Aerts S., A Single-Cell Transcriptome Atlas of the Aging Drosophila Brain. Cell. 174, 982–998.e20 (2018). - PMC - PubMed
-
- Tabula Sapiens Consortium*, Jones R. C., Karkanias J., Krasnow M. A., Pisco A. O., Quake S. R., Salzman J., Yosef N., Bulthaup B., Brown P., Harper W., Hemenez M., Ponnusamy R., Salehi A., Sanagavarapu B. A., Spallino E., Aaron K. A., Concepcion W., Gardner J. M., Kelly B., Neidlinger N., Wang Z., Crasta S., Kolluru S., Morri M., Pisco A. O., Tan S. Y., Travaglini K. J., Xu C., Alcántara-Hernández M., Almanzar N., Antony J., Beyersdorf B., Burhan D., Calcuttawala K., Carter M. M., Chan C. K. F., Chang C. A., Chang S., Colville A., Crasta S., Culver R. N., Cvijović I., D’Amato G., Ezran C., Galdos F. X., Gillich A., Goodyer W. R., Hang Y., Hayashi A., Houshdaran S., Huang X., Irwin J. C., Jang S., Juanico J. V., Kershner A. M., Kim S., Kiss B., Kolluru S., Kong W., Kumar M. E., Kuo A. H., Leylek R., Li B., Loeb G. B., Lu W.-J., Mantri S., Markovic M., McAlpine P. L., de Morree A., Morri M., Mrouj K., Mukherjee S., Muser T., Neuhöfer P., Nguyen T. D., Perez K., Phansalkar R., Pisco A. O., Puluca N., Qi Z., Rao P., Raquer-McKay H., Schaum N., Scott B., Seddighzadeh B., Segal J., Sen S., Sikandar S., Spencer S. P., Steffes L. C., Subramaniam V. R., Swarup A., Swift M., Travaglini K. J., Van Treuren W., Trimm E., Veizades S., Vijayakumar S., Vo K. C., Vorperian S. K., Wang W., Weinstein H. N. W., Winkler J., Wu T. T. H., Xie J., Yung A. R., Zhang Y., Detweiler A. M., Mekonen H., Neff N. F., Sit R. V., Tan M., Yan J., Bean G. R., Charu V., Forgó E., Martin B. A., Ozawa M. G., Silva O., Tan S. Y., Toland A., Vemuri V. N. P., Afik S., Awayan K., Botvinnik O. B., Byrne A., Chen M., Dehghannasiri R., Detweiler A. M., Gayoso A., Granados A. A., Li Q., Mahmoudabadi G., McGeever A., de Morree A., Olivieri J. E., Park M., Pisco A. O., Ravikumar N., Salzman J., Stanley G., Swift M., Tan M., Tan W., Tarashansky A. J., Vanheusden R., Vorperian S. K., Wang P., Wang S., Xing G., Xu C., Yosef N., Alcántara-Hernández M., Antony J., Chan C. K. F., Chang C. A., Colville A., Crasta S., Culver R., Dethlefsen L., Ezran C., Gillich A., Hang Y., Ho P.-Y., Irwin J. C., Jang S., Kershner A. M., Kong W., Kumar M. E., Kuo A. H., Leylek R., Liu S., Loeb G. B., Lu W.-J., Maltzman J. S., Metzger R. J., de Morree A., Neuhöfer P., Perez K., Phansalkar R., Qi Z., Rao P., Raquer-McKay H., Sasagawa K., Scott B., Sinha R., Song H., Spencer S. P., Swarup A., Swift M., Travaglini K. J., Trimm E., Veizades S., Vijayakumar S., Wang B., Wang W., Winkler J., Xie J., Yung A. R., Artandi S. E., Beachy P. A., Clarke M. F., Giudice L. C., Huang F. W., Huang K. C., Idoyaga J., Kim S. K., Krasnow M., Kuo C. S., Nguyen P., Quake S. R., Rando T. A., Red-Horse K., Reiter J., Relman D. A., Sonnenburg J. L., Wang B., Wu A., Wu S. M., Wyss-Coray T., The Tabula Sapiens: A multiple-organ, single-cell transcriptomic atlas of humans. Science. 376, eabl4896 (2022). - PMC - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources