

This is a preprint.
Erg251 has complex and pleiotropic effects on azole susceptibility, filamentation, and stress response phenotypes
- PMID: 38496635
- PMCID: PMC10942443
- DOI: 10.1101/2024.03.06.583770
Erg251 has complex and pleiotropic effects on azole susceptibility, filamentation, and stress response phenotypes
Update in
-
Erg251 has complex and pleiotropic effects on sterol composition, azole susceptibility, filamentation, and stress response phenotypes.PLoS Pathog. 2024 Jul 30;20(7):e1012389. doi: 10.1371/journal.ppat.1012389. eCollection 2024 Jul. PLoS Pathog. 2024. PMID: 39078851 Free PMC article.
Abstract
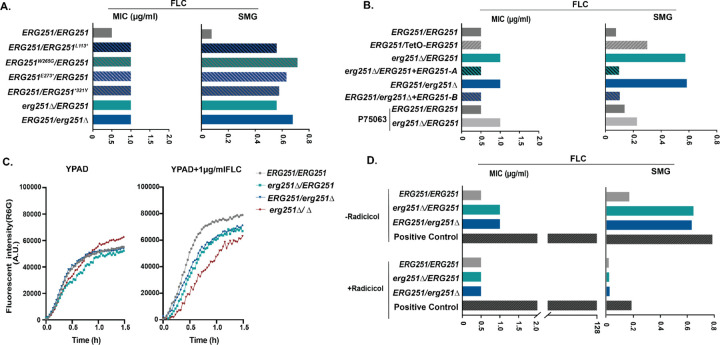
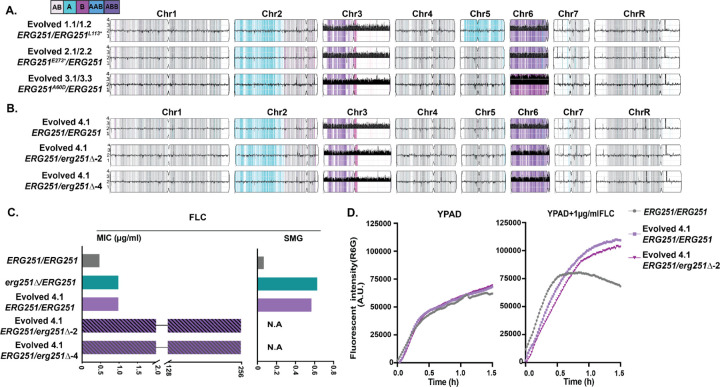
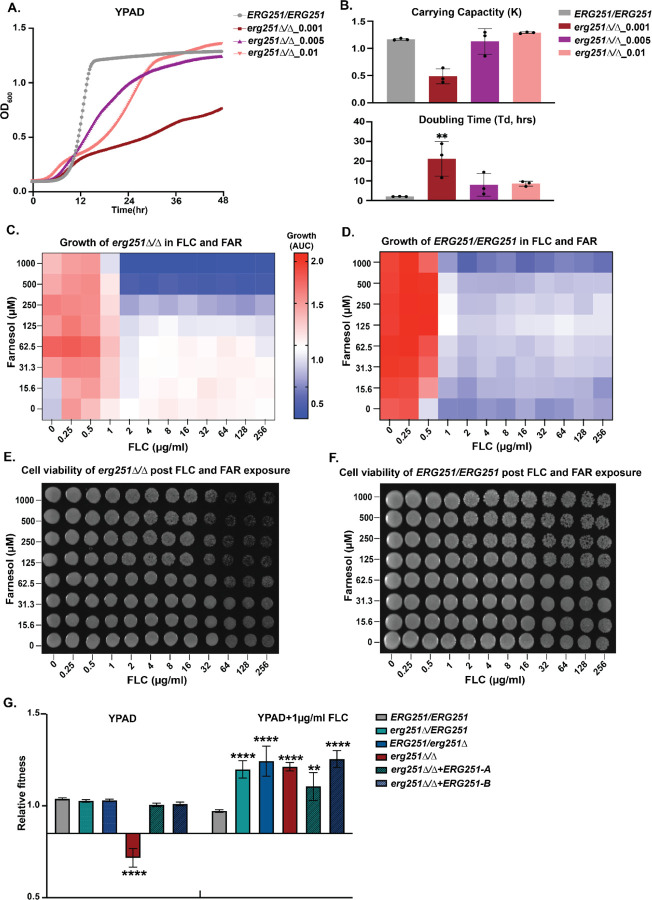
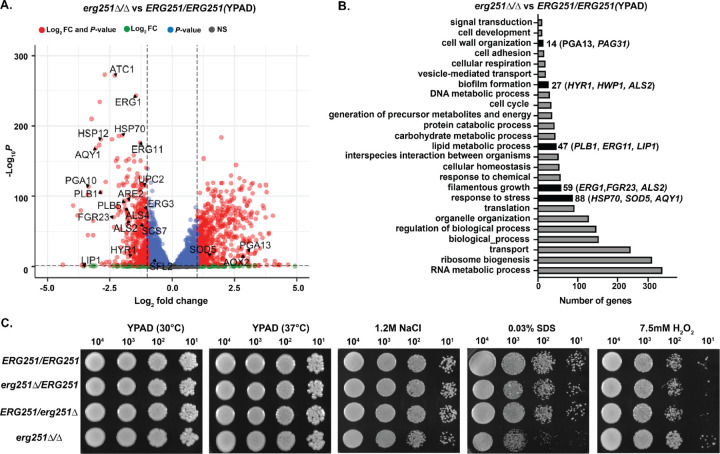
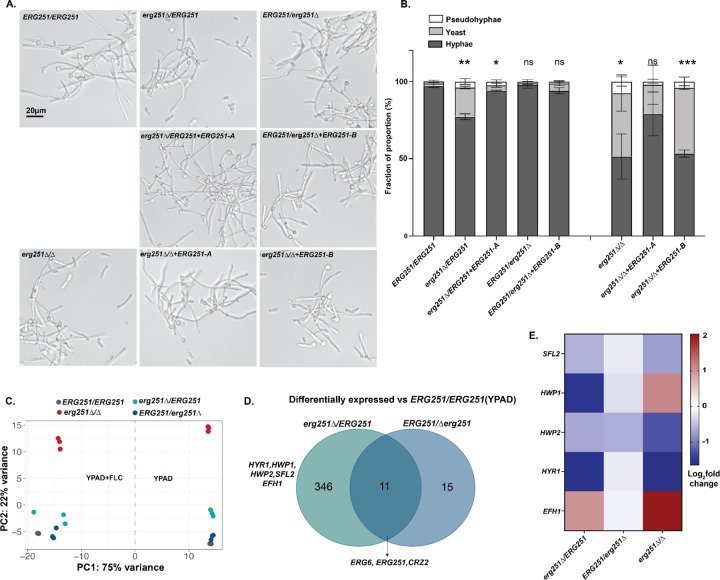
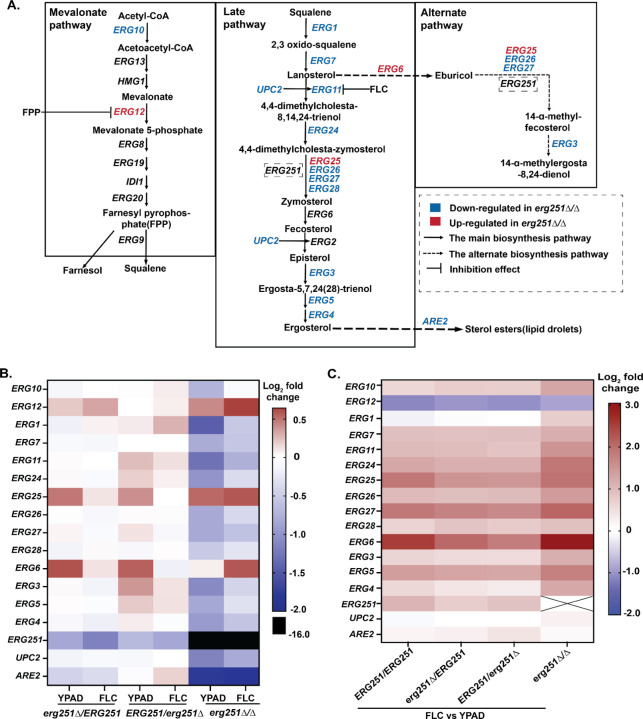
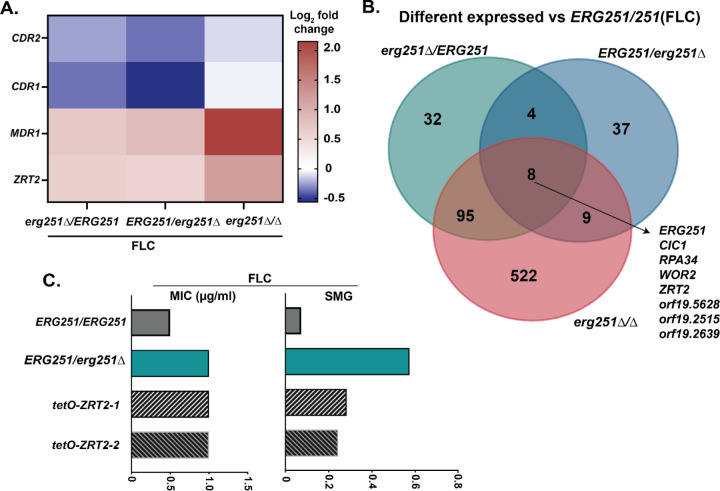
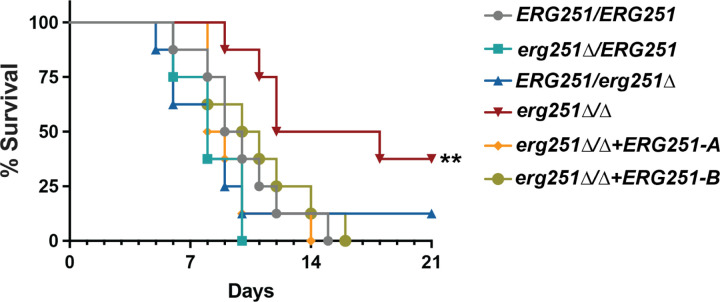
Ergosterol is essential for fungal cell membrane integrity and growth, and numerous antifungal drugs target ergosterol. Inactivation or modification of ergosterol biosynthetic genes can lead to changes in antifungal drug susceptibility, filamentation and stress response. Here, we found that the ergosterol biosynthesis gene ERG251 is a hotspot for point mutations during adaptation to antifungal drug stress within two distinct genetic backgrounds of Candida albicans. Heterozygous point mutations led to single allele dysfunction of ERG251 and resulted in azole tolerance in both genetic backgrounds. This is the first known example of point mutations causing azole tolerance in C. albicans. Importantly, single allele dysfunction of ERG251 in combination with recurrent chromosome aneuploidies resulted in bona fide azole resistance. Homozygous deletions of ERG251 caused increased fitness in low concentrations of fluconazole and decreased fitness in rich medium, especially at low initial cell density. Dysfunction of ERG251 resulted in transcriptional upregulation of the alternate sterol biosynthesis pathway and ZRT2, a Zinc transporter. Notably, we determined that overexpression of ZRT2 is sufficient to increase azole tolerance in C. albicans. Our combined transcriptional and phenotypic analyses revealed the pleiotropic effects of ERG251 on stress responses including cell wall, osmotic and oxidative stress. Interestingly, while loss of either allele of ERG251 resulted in similar antifungal drug responses, we observed functional divergence in filamentation regulation between the two alleles of ERG251 (ERG251-A and ERG251-B) with ERG251-A exhibiting a dominant role in the SC5314 genetic background. Finally, in a murine model of systemic infection, homozygous deletion of ERG251 resulted in decreased virulence while the heterozygous deletion mutants maintain their pathogenicity. Overall, this study provides extensive genetic, transcriptional and phenotypic analysis for the effects of ERG251 on drug susceptibility, fitness, filamentation and stress responses.
Figures
Similar articles
-
Erg251 has complex and pleiotropic effects on sterol composition, azole susceptibility, filamentation, and stress response phenotypes.PLoS Pathog. 2024 Jul 30;20(7):e1012389. doi: 10.1371/journal.ppat.1012389. eCollection 2024 Jul. PLoS Pathog. 2024. PMID: 39078851 Free PMC article.
-
Azole resistance by loss of function of the sterol Δ⁵,⁶-desaturase gene (ERG3) in Candida albicans does not necessarily decrease virulence.Antimicrob Agents Chemother. 2012 Apr;56(4):1960-8. doi: 10.1128/AAC.05720-11. Epub 2012 Jan 17. Antimicrob Agents Chemother. 2012. PMID: 22252807 Free PMC article.
-
Genetic Analysis of Candida auris Implicates Hsp90 in Morphogenesis and Azole Tolerance and Cdr1 in Azole Resistance.mBio. 2019 Jan 29;10(1):e02529-18. doi: 10.1128/mBio.02529-18. mBio. 2019. PMID: 30696744 Free PMC article.
-
Antifungal drug resistance in pathogenic fungi.Med Mycol. 1998;36 Suppl 1:119-28. Med Mycol. 1998. PMID: 9988500 Review.
-
Resistance in human pathogenic yeasts and filamentous fungi: prevalence, underlying molecular mechanisms and link to the use of antifungals in humans and the environment.Dan Med J. 2016 Oct;63(10):B5288. Dan Med J. 2016. PMID: 27697142 Review.
References
-
- Pfaller MA. Antifungal drug resistance: mechanisms, epidemiology, and consequences for treatment. Am J Med. 2012;125: S3–13. - PubMed
-
- Perea S, López-Ribot JL, Kirkpatrick WR, McAtee RK, Santillán RA, Martínez M, et al. Prevalence of molecular mechanisms of resistance to azole antifungal agents in Candida albicans strains displaying high-level fluconazole resistance isolated from human immunodeficiency virus-infected patients. Antimicrob Agents Chemother. 2001;45: 2676–2684. - PMC - PubMed
-
- Andes DR, Safdar N, Baddley JW, Playford G, Reboli AC, Rex JH, et al. Impact of treatment strategy on outcomes in patients with candidemia and other forms of invasive candidiasis: a patient-level quantitative review of randomized trials. Clin Infect Dis. 2012;54: 1110–1122. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources