

This is a preprint.
Crimean-Congo Hemorrhagic Fever Survivors Elicit Protective Non-Neutralizing Antibodies that Target 11 Overlapping Regions on Viral Glycoprotein GP38
- PMID: 38496658
- PMCID: PMC10942344
- DOI: 10.1101/2024.03.02.583110
Crimean-Congo Hemorrhagic Fever Survivors Elicit Protective Non-Neutralizing Antibodies that Target 11 Overlapping Regions on Viral Glycoprotein GP38
Update in
-
Crimean-Congo hemorrhagic fever survivors elicit protective non-neutralizing antibodies that target 11 overlapping regions on glycoprotein GP38.Cell Rep. 2024 Jul 23;43(7):114502. doi: 10.1016/j.celrep.2024.114502. Epub 2024 Jul 13. Cell Rep. 2024. PMID: 39002130 Free PMC article.
Abstract
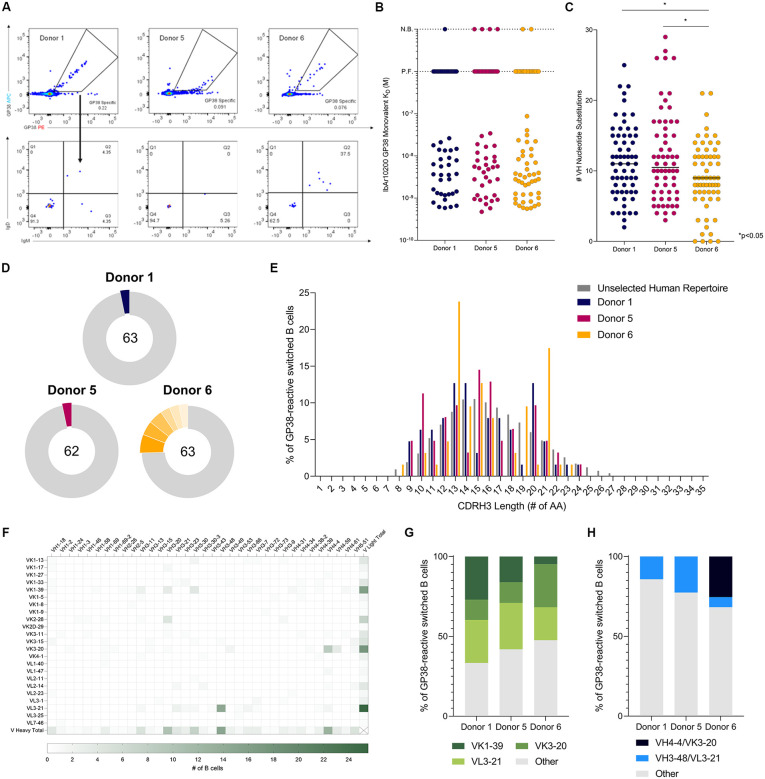
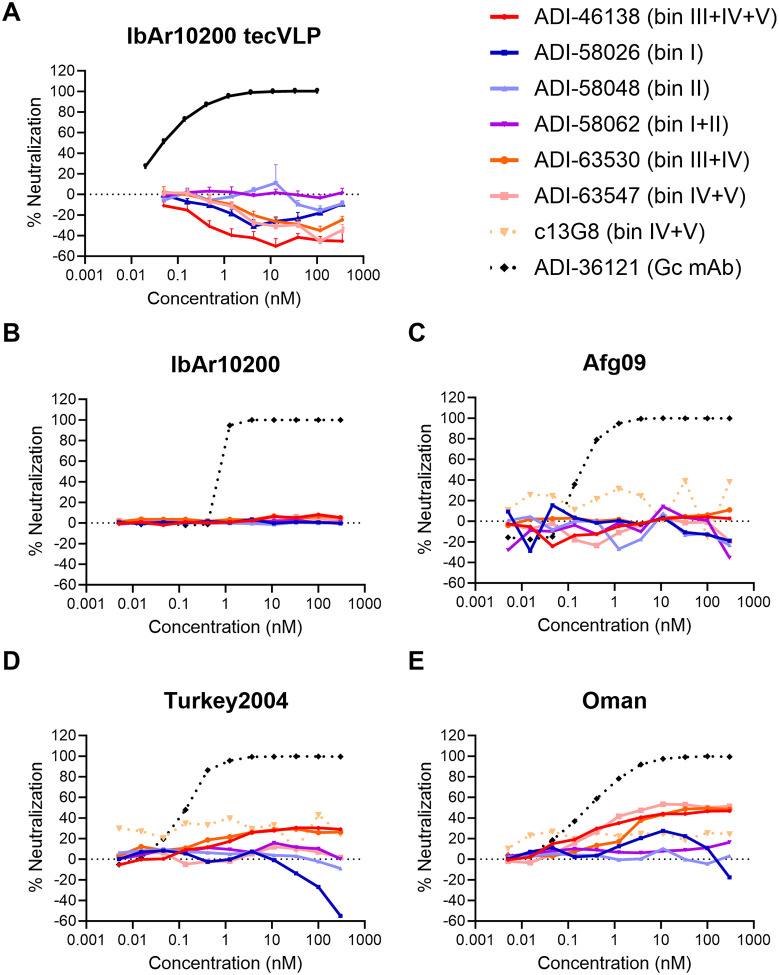
Crimean-Congo hemorrhagic fever virus can cause lethal disease in humans yet there are no approved medical countermeasures. Viral glycoprotein GP38, unique to Nairoviridae, is a target of protective antibodies, but extensive mapping of the human antibody response to GP38 has not been previously performed. Here, we isolated 188 GP38-specific antibodies from human survivors of infection. Competition experiments showed that these antibodies bind across five distinct antigenic sites, encompassing eleven overlapping regions. Additionally, we reveal structures of GP38 bound with nine of these antibodies targeting different antigenic sites. Although GP38-specific antibodies were non-neutralizing, several antibodies were found to have protection equal to or better than murine antibody 13G8 in two highly stringent rodent models of infection. Together, these data expand our understanding regarding this important viral protein and inform the development of broadly effective CCHFV antibody therapeutics.
Keywords: CCHFV; Crimean-Congo hemorrhagic fever virus; GP38; Nairoviridae; antibody therapeutics; human monoclonal antibody; tickborne; viral glycoprotein.
Conflict of interest statement
DECLARATION OF INTERESTS N.T.P., E.C., and J.L. are employees and shareholders of Adimab, LLC. D.P.M., L.M.W., O.S.S., V.H., and M.D. are shareholders of Adimab.
Figures





Similar articles
-
Crimean-Congo hemorrhagic fever survivors elicit protective non-neutralizing antibodies that target 11 overlapping regions on glycoprotein GP38.Cell Rep. 2024 Jul 23;43(7):114502. doi: 10.1016/j.celrep.2024.114502. Epub 2024 Jul 13. Cell Rep. 2024. PMID: 39002130 Free PMC article.
-
Structure and Characterization of Crimean-Congo Hemorrhagic Fever Virus GP38.J Virol. 2020 Mar 31;94(8):e02005-19. doi: 10.1128/JVI.02005-19. Print 2020 Mar 31. J Virol. 2020. PMID: 31996434 Free PMC article.
-
Structural characterization of protective non-neutralizing antibodies targeting Crimean-Congo hemorrhagic fever virus.Nat Commun. 2022 Nov 26;13(1):7298. doi: 10.1038/s41467-022-34923-0. Nat Commun. 2022. PMID: 36435827 Free PMC article.
-
Crimean-Congo Hemorrhagic Fever Virus for Clinicians-Diagnosis, Clinical Management, and Therapeutics.Emerg Infect Dis. 2024 May;30(5):864-873. doi: 10.3201/eid3005.231648. Emerg Infect Dis. 2024. PMID: 38666553 Free PMC article. Review.
-
Crimean-Congo Hemorrhagic Fever.Lab Med. 2015 Summer;46(3):180-9. doi: 10.1309/LMN1P2FRZ7BKZSCO. Lab Med. 2015. PMID: 26199256 Review.
References
-
- Spengler J.R., Estrada-Peña A., Garrison A.R., Schmaljohn C., Spiropoulou C.F., Bergeron É., and Bente D.A. (2016). A chronological review of experimental infection studies of the role of wild animals and livestock in the maintenance and transmission of Crimean-Congo hemorrhagic fever virus. Antiviral Res 135, 31–47. 10.1016/j.antiviral.2016.09.013. - DOI - PMC - PubMed
-
- Nurettin C., Engin B., Sukru T., Munir A., Zati V., and Aykut O. (2022). The Seroprevalence of Crimean-Congo Hemorrhagic Fever in Wild and Domestic Animals: An Epidemiological Update for Domestic Animals and First Seroevidence in Wild Animals from Turkiye. Vet Sci 9. 10.3390/vetsci9090462. - DOI - PMC - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources