

The dynamics of the flavin, NADPH, and active site loops determine the mechanism of activation of class B flavin-dependent monooxygenases
- PMID: 38501462
- PMCID: PMC10962481
- DOI: 10.1002/pro.4935
The dynamics of the flavin, NADPH, and active site loops determine the mechanism of activation of class B flavin-dependent monooxygenases
Abstract
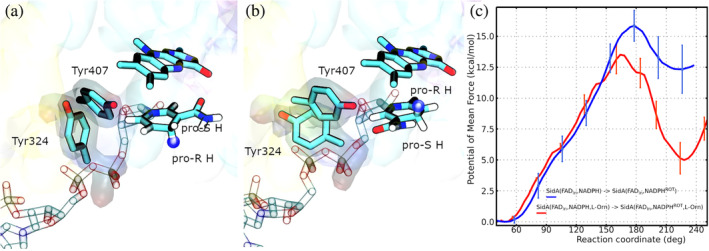
Flavin-dependent monooxygenases (FMOs) constitute a diverse enzyme family that catalyzes crucial hydroxylation, epoxidation, and Baeyer-Villiger reactions across various metabolic pathways in all domains of life. Due to the intricate nature of this enzyme family's mechanisms, some aspects of their functioning remain unknown. Here, we present the results of molecular dynamics computations, supplemented by a bioinformatics analysis, that clarify the early stages of their catalytic cycle. We have elucidated the intricate binding mechanism of NADPH and L-Orn to a class B monooxygenase, the ornithine hydroxylase from known as SidA. Our investigation involved a comprehensive characterization of the conformational changes associated with the FAD (Flavin Adenine Dinucleotide) cofactor, transitioning from the out to the in position. Furthermore, we explored the rotational dynamics of the nicotinamide ring of NADPH, shedding light on its role in facilitating FAD reduction, supported by experimental evidence. Finally, we also analyzed the extent of conservation of two Tyr-loops that play critical roles in the process.
Keywords: NADPH binding; NADPH dynamics; flavin dynamics; flavin-dependent monooxygenases; ornithine binding; uncoupling.
© 2024 The Protein Society.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures








Similar articles
-
Trapping conformational states of a flavin-dependent N-monooxygenase in crystallo reveals protein and flavin dynamics.J Biol Chem. 2020 Sep 18;295(38):13239-13249. doi: 10.1074/jbc.RA120.014750. Epub 2020 Jul 28. J Biol Chem. 2020. PMID: 32723870 Free PMC article.
-
Structural Determinants of Flavin Dynamics in a Class B Monooxygenase.Biochemistry. 2020 Dec 8;59(48):4609-4616. doi: 10.1021/acs.biochem.0c00783. Epub 2020 Nov 23. Biochemistry. 2020. PMID: 33226785
-
Two structures of an N-hydroxylating flavoprotein monooxygenase: ornithine hydroxylase from Pseudomonas aeruginosa.J Biol Chem. 2011 Sep 9;286(36):31789-98. doi: 10.1074/jbc.M111.265876. Epub 2011 Jul 13. J Biol Chem. 2011. PMID: 21757711 Free PMC article.
-
The devil is in the details: The chemical basis and mechanistic versatility of flavoprotein monooxygenases.Arch Biochem Biophys. 2021 Feb 15;698:108732. doi: 10.1016/j.abb.2020.108732. Epub 2020 Dec 24. Arch Biochem Biophys. 2021. PMID: 33358998 Review.
-
Monooxygenation of aromatic compounds by flavin-dependent monooxygenases.Protein Sci. 2019 Jan;28(1):8-29. doi: 10.1002/pro.3525. Protein Sci. 2019. PMID: 30311986 Free PMC article. Review.
Cited by
-
Enhancing Bacillus cereus antibacterial ability through improved cofactor supply.Microb Cell Fact. 2025 Mar 6;24(1):52. doi: 10.1186/s12934-025-02666-4. Microb Cell Fact. 2025. PMID: 40050842 Free PMC article.
References
-
- Badieyan S, Bach RD, Sobrado P. Mechanism of N‐hydroxylation catalyzed by flavin‐dependent monooxygenases. J Org Chem. 2015;80:2139–2147. - PubMed
-
- Beaty NB, Ballou DP. The reductive half‐reaction of liver microsomal FAD‐containing monooxygenase. J Biol Chem. 1981;256:4611–4618. - PubMed
-
- Campbell AC, Robinson R, Mena‐Aguilar D, Sobrado P, Tanner JJ. Structural determinants of flavin dynamics in a class B monooxygenase. Biochemistry. 2020;59:4609–4616. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
