

Irgm proteins attenuate inflammatory disease in mouse models of genital Chlamydia infection
- PMID: 38501887
- PMCID: PMC11005385
- DOI: 10.1128/mbio.00303-24
Irgm proteins attenuate inflammatory disease in mouse models of genital Chlamydia infection
Abstract
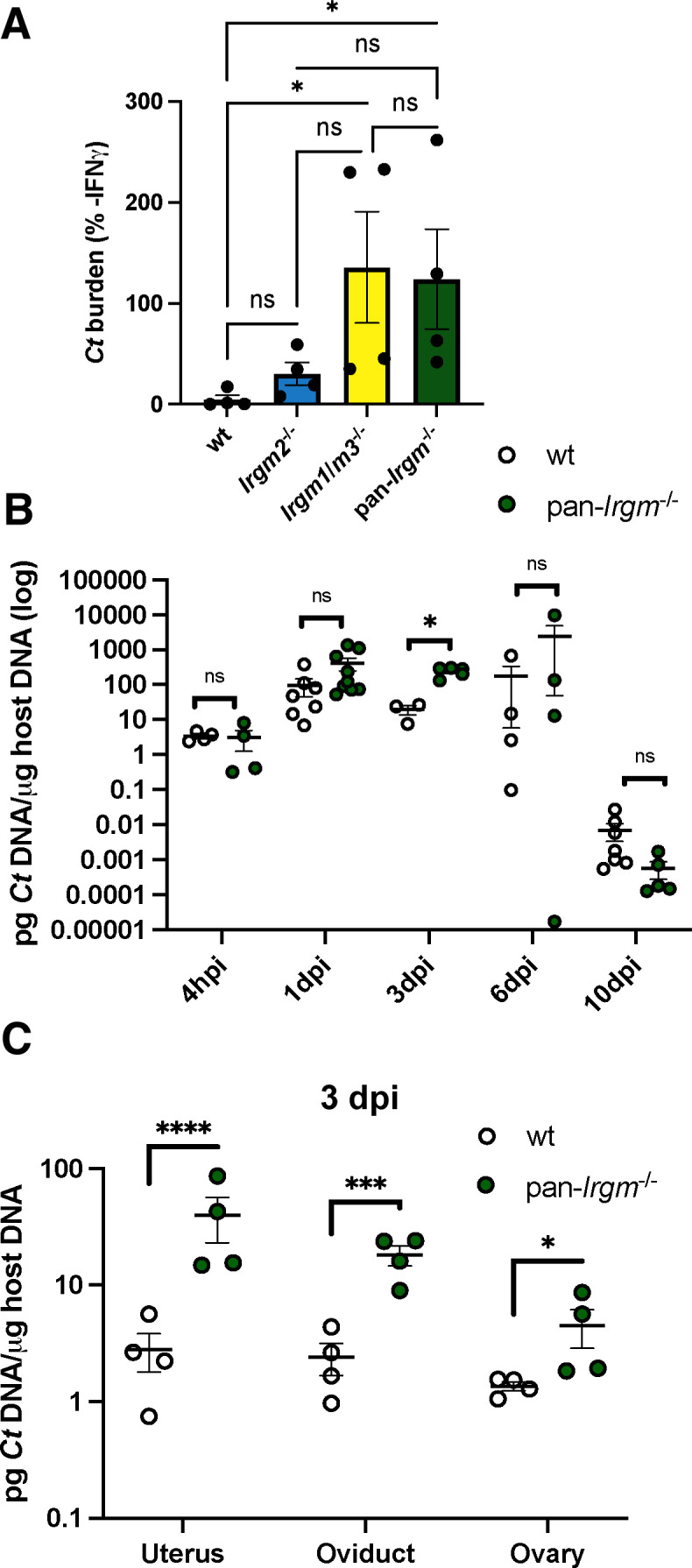
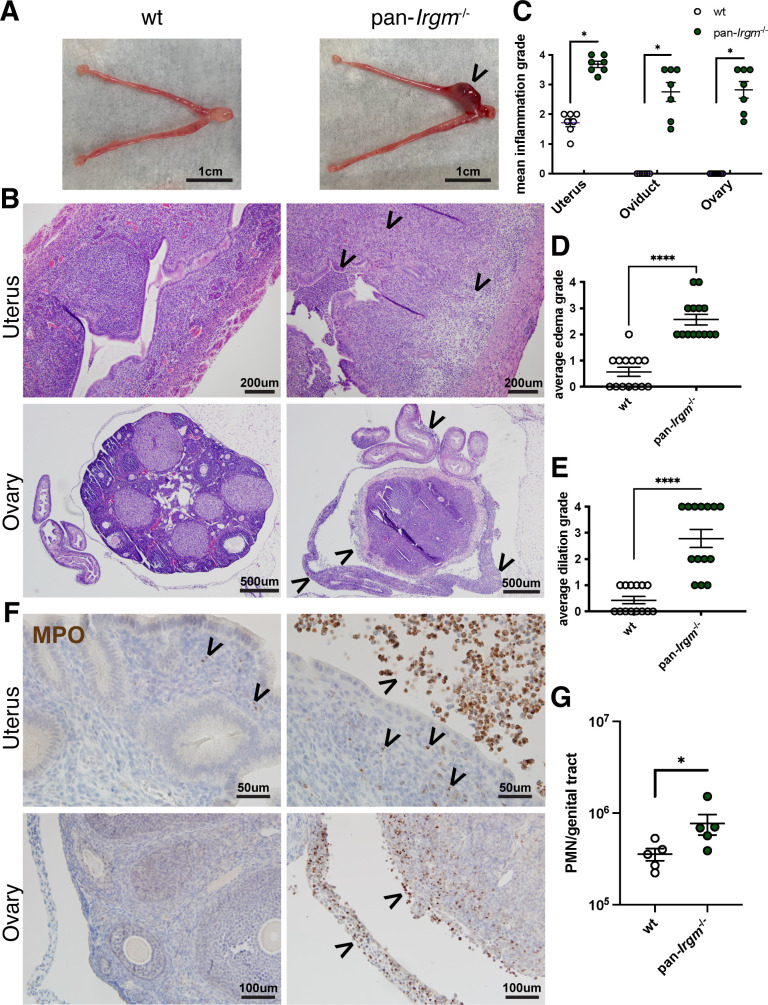
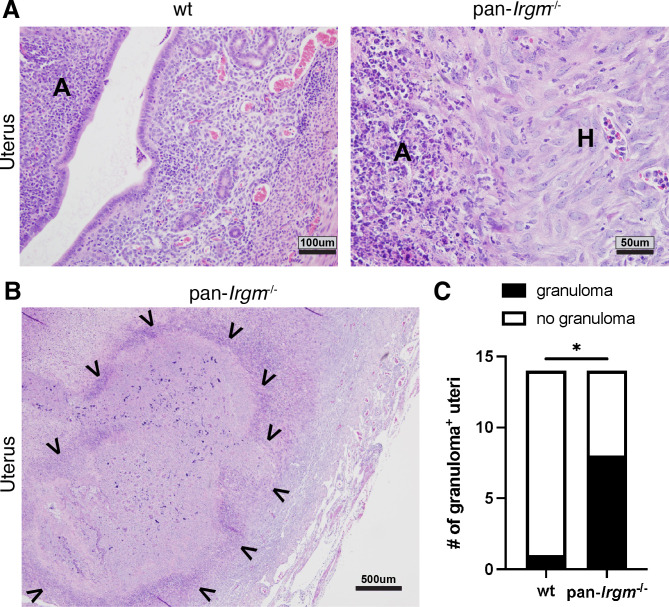
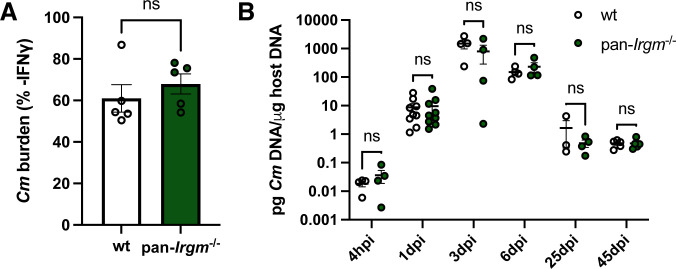
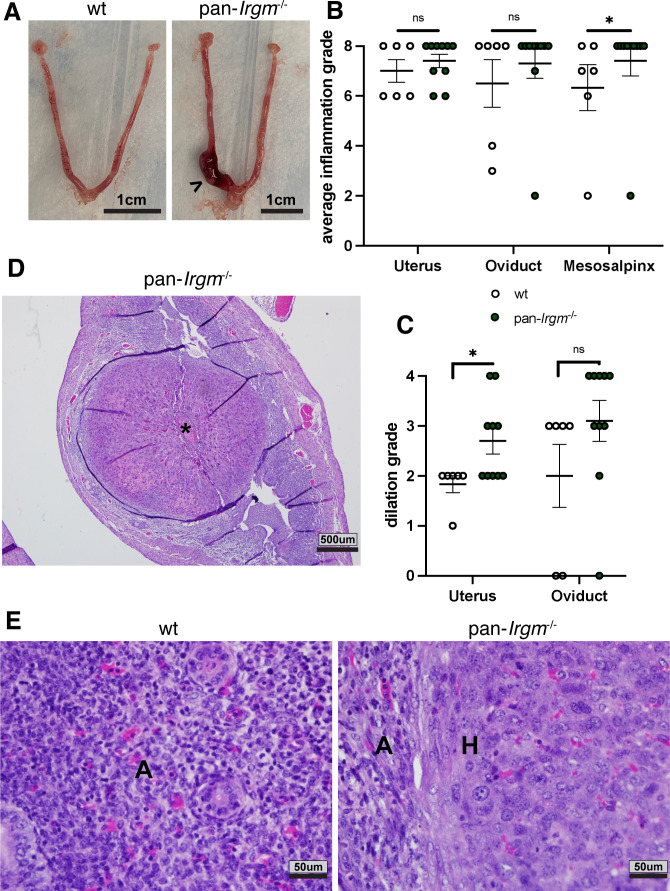
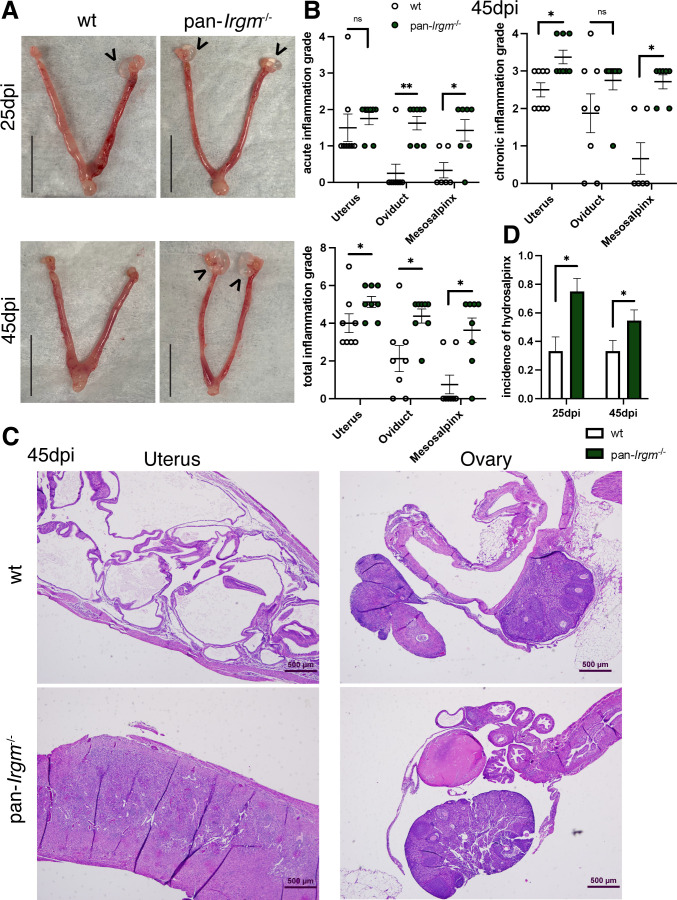
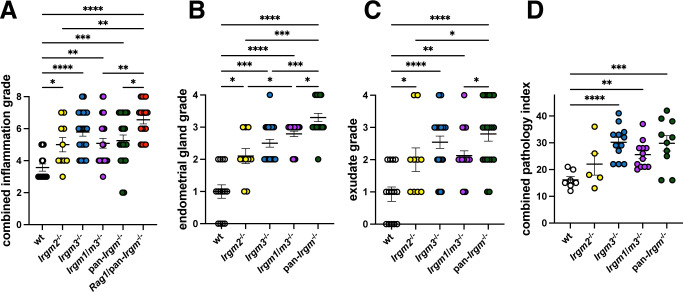
Chlamydiae are obligate intracellular bacterial pathogens that may cause genital pathology via induction of destructive host immune responses. Human-adapted Chlamydia trachomatis causes inflammatory disease in human hosts but is easily cleared in mice, and mouse-adapted Chlamydia muridarum establishes a productive and pathogenic infection in murine hosts. While numerous anti-chlamydial host resistance factors have been discovered in mice and humans alike, little is known about host factors promoting host fitness independent of host resistance. Here, we show that interferon-inducible immunity-related GTPase M (Irgm) proteins function as such host factors ameliorating infection-associated sequalae in the murine female genital tract, thus characterizing Irgm proteins as mediators of disease tolerance. Specifically, we demonstrate that mice deficient for all three murine Irgm paralogs (pan-Irgm-/-) are defective for cell-autonomous immunity to C. trachomatis, which correlates with an early and transient increase in bacterial burden and sustained hyperinflammation in vivo. In contrast, upon infection of pan-Irgm-/- mice with C. muridarum, bacterial burden is unaffected, yet genital inflammation and scarring pathology are nonetheless increased, demonstrating that Irgm proteins can promote host fitness without altering bacterial burden. Additionally, pan-Irgm-/- mice display increased granulomatous inflammation in genital Chlamydia infection, implicating Irgm proteins in the regulation of granuloma formation and maintenance. These findings demonstrate that Irgm proteins regulate pathogenic immune responses to Chlamydia infection in vivo, establishing an effective infection model to examine the immunoregulatory functions and mechanisms of Irgm proteins.
Importance: In response to genital Chlamydia infection, the immune system mounts a proinflammatory response to resist the pathogen, yet inflammation must be tightly controlled to avoid collateral damage and scarring to host genital tissue. Variation in the human IRGM gene is associated with susceptibility to autoinflammatory diseases but its role in ameliorating inflammatory diseases caused by infections is poorly defined. Here, we use mice deficient for all three murine Irgm paralogs to demonstrate that Irgm proteins not only provide host resistance to Chlamydia infections but also limit associated inflammation in the female genital tract. In particular, we find that murine Irgm expression prevents granulomatous inflammation, which parallels inflammatory diseases associated with variants in human IRGM. Our findings therefore establish genital Chlamydia infection as a useful model to study the roles for Irgm proteins in both promoting protective immunity and limiting pathogenic inflammation.
Keywords: Chlamydia; IRGM; disease tolerance; immunity-related GTPases; immunopathology; interferons; sexually transmitted diseases.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Braxton JDD, Emerson B. 2018. Sexually transmitted surveillance 2017. Centers for disease control and prevention
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
