

Structural insight into Escherichia coli CsgA amyloid fibril assembly
- PMID: 38501920
- PMCID: PMC11005368
- DOI: 10.1128/mbio.00419-24
Structural insight into Escherichia coli CsgA amyloid fibril assembly
Abstract
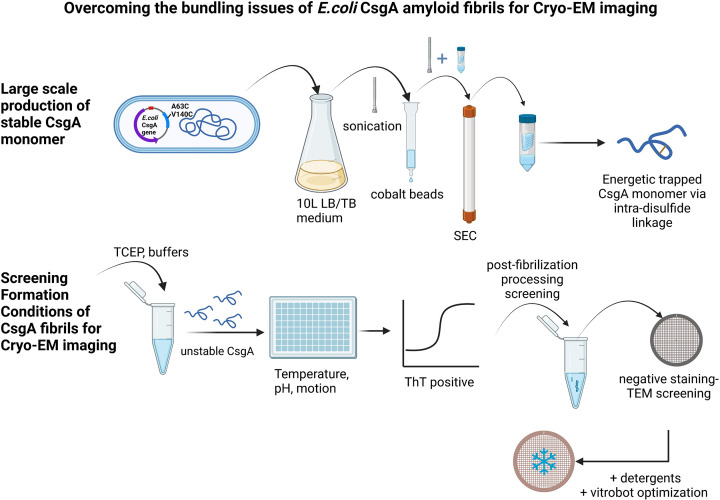
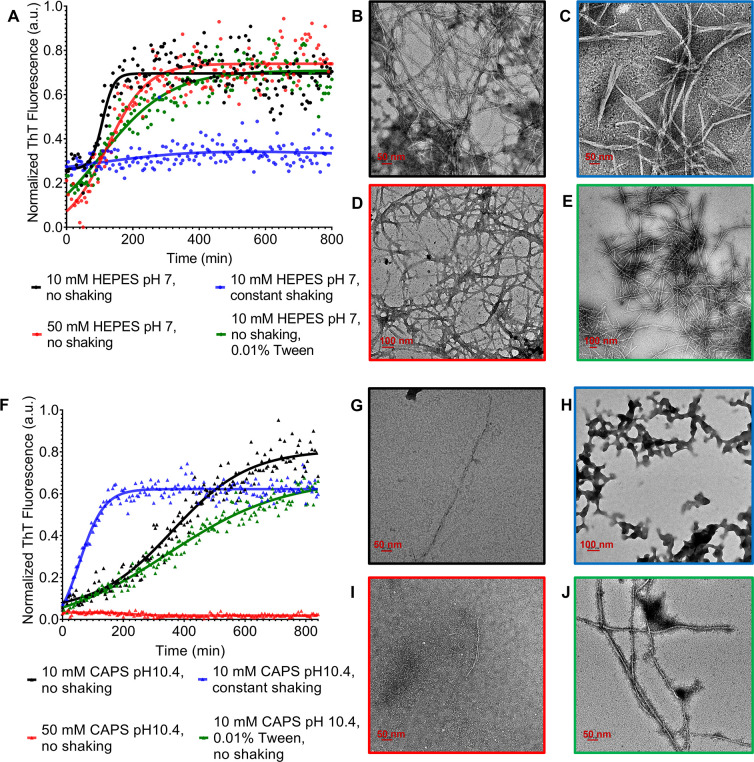
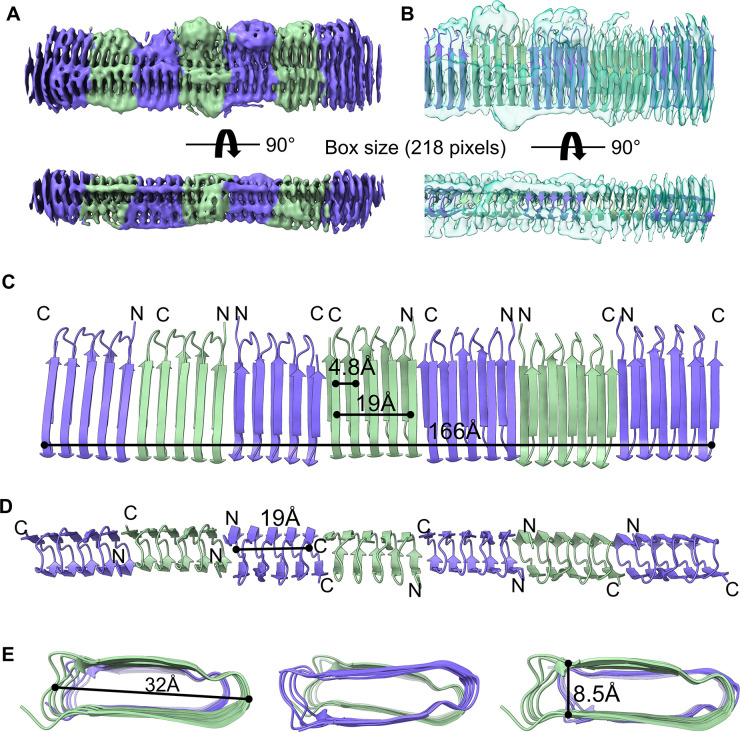
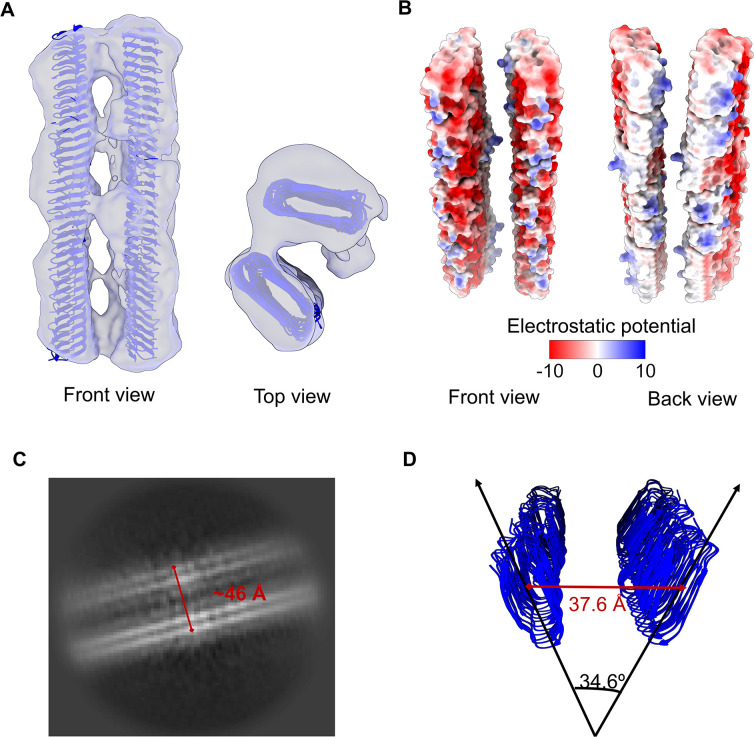
The discovery of functional amyloids in bacteria dates back several decades, and our understanding of the Escherichia coli curli biogenesis system has gradually expanded over time. However, due to its high aggregation propensity and intrinsically disordered nature, CsgA, the main structural component of curli fibrils, has eluded comprehensive structural characterization. Recent advancements in cryo-electron microscopy (cryo-EM) offer a promising tool to achieve high-resolution structural insights into E. coli CsgA fibrils. In this study, we outline an approach to addressing the colloidal instability challenges associated with CsgA, achieved through engineering and electrostatic repulsion. Then, we present the cryo-EM structure of CsgA fibrils at 3.62 Å resolution. This structure provides new insights into the cross-β structure of E. coli CsgA. Additionally, our study identifies two distinct spatial arrangements within several CsgA fibrils, a 2-CsgA-fibril pair and a 3-CsgA-fibril bundle, shedding light on the intricate hierarchy of the biofilm extracellular matrix and laying the foundation for precise manipulation of CsgA-derived biomaterials.IMPORTANCEThe visualization of the architecture of Escherichia coli CsgA amyloid fibril has been a longstanding research question, for which a high-resolution structure is still unavailable. CsgA serves as a major subunit of curli, the primary component of the extracellular matrix generated by bacteria. The support provided by this extracellular matrix enables bacterial biofilms to resist antibiotic treatment, significantly impacting human health. CsgA has been identified in members of Enterobacteriaceae, with pathogenic E. coli being the most well-known model system. Our novel insights into the structure of E. coli CsgA protofilaments form the basis for drug design targeting diseases associated with biofilms. Additionally, CsgA is widely researched in biomaterials due to its self-assembly characteristics. The resolved spatial arrangements of CsgA amyloids revealed in our study will further enhance the precision design of functional biomaterials. Therefore, our study uniquely contributes to the understanding of CsgA amyloids for both microbiology and material science.
Keywords: bacteria; biofilms; curli; electron microscopy; functional amyloids; protein aggregation; protein structure-function.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Gallat F-X, Laganowsky A, Wood K, Gabel F, van Eijck L, Wuttke J, Moulin M, Härtlein M, Eisenberg D, Colletier J-P, Zaccai G, Weik M. 2012. Dynamical coupling of intrinsically disordered proteins and their hydration water: comparison with folded soluble and membrane proteins. Biophys J 103:129–136. doi: 10.1016/j.bpj.2012.05.027 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
