

Contrasting somatic mutation patterns in aging human neurons and oligodendrocytes
- PMID: 38503282
- PMCID: PMC11062076
- DOI: 10.1016/j.cell.2024.02.025
Contrasting somatic mutation patterns in aging human neurons and oligodendrocytes
Abstract
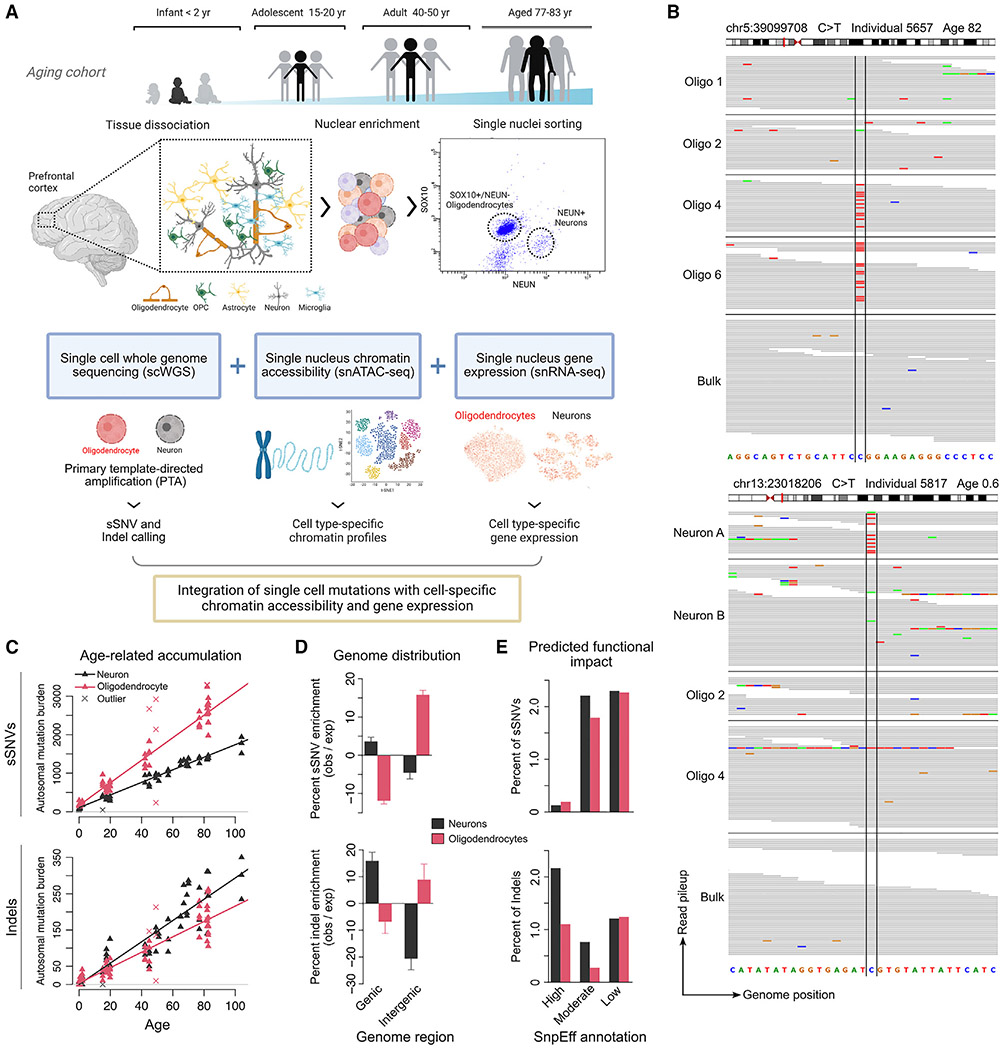
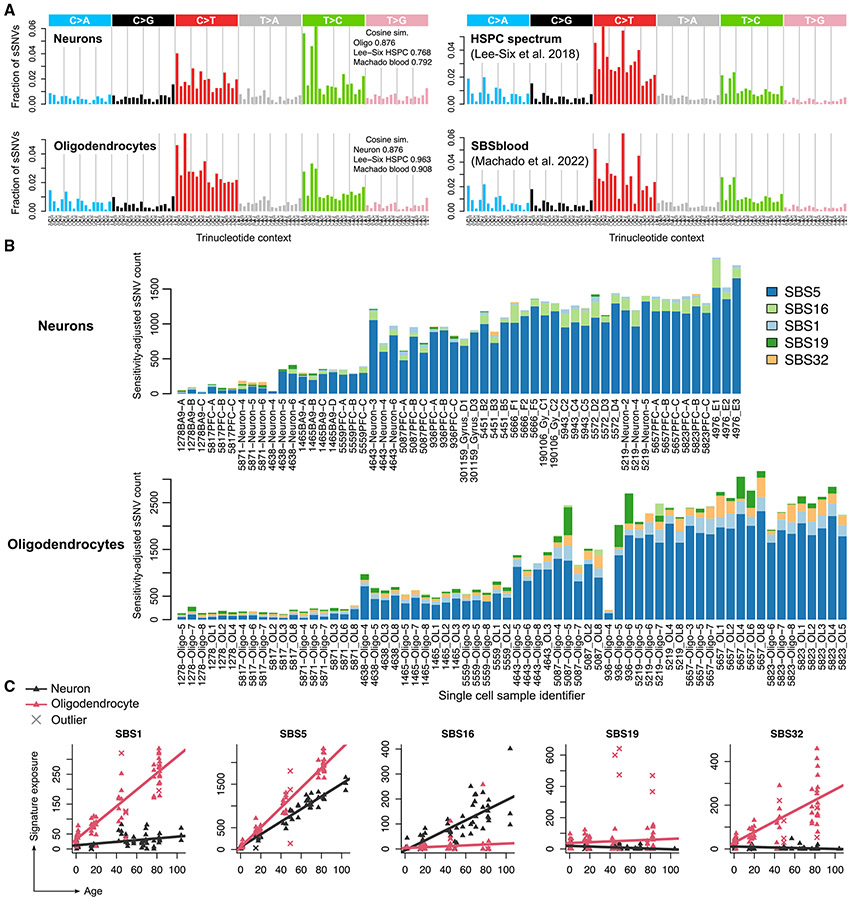
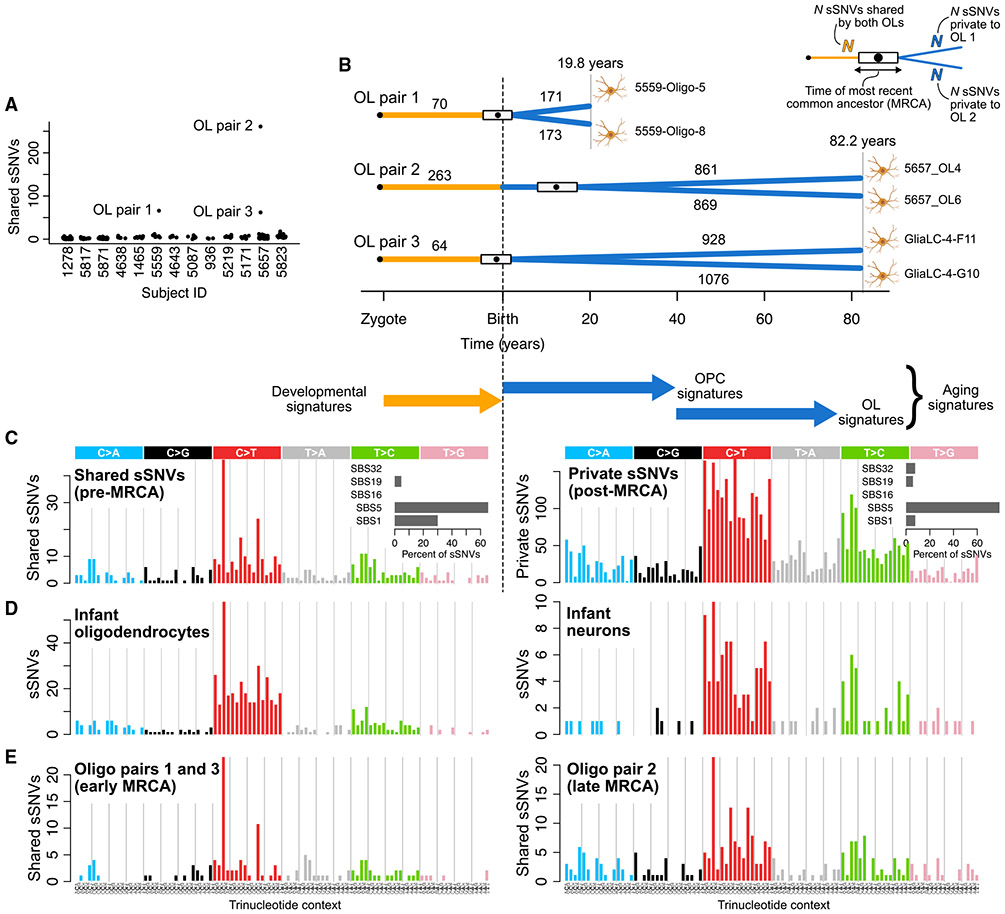
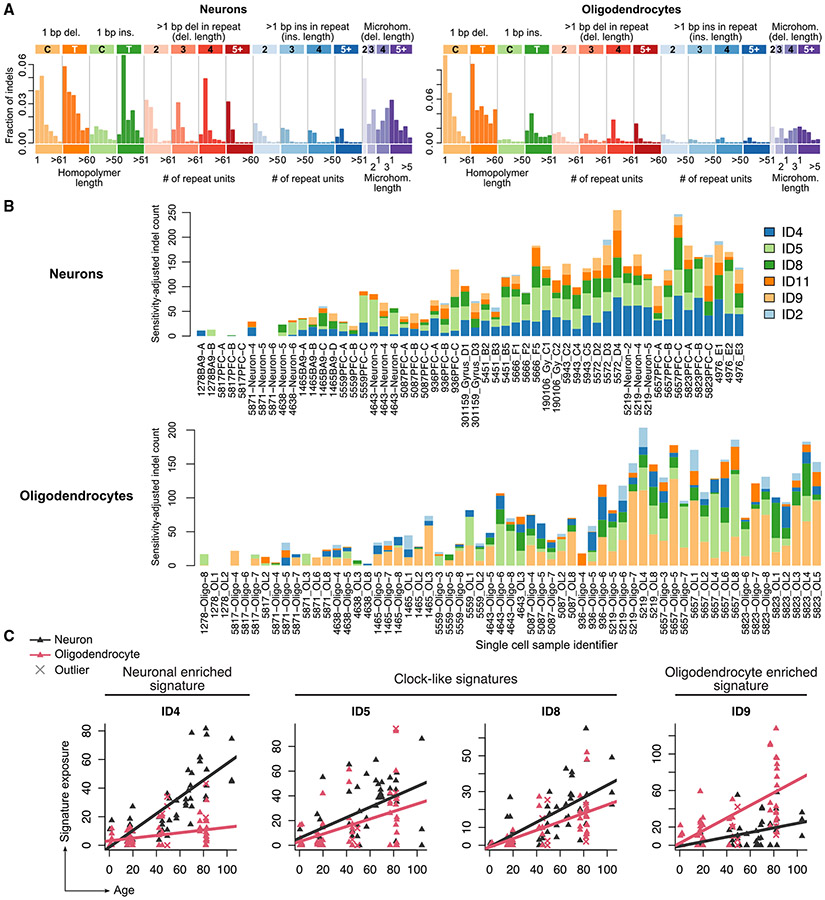
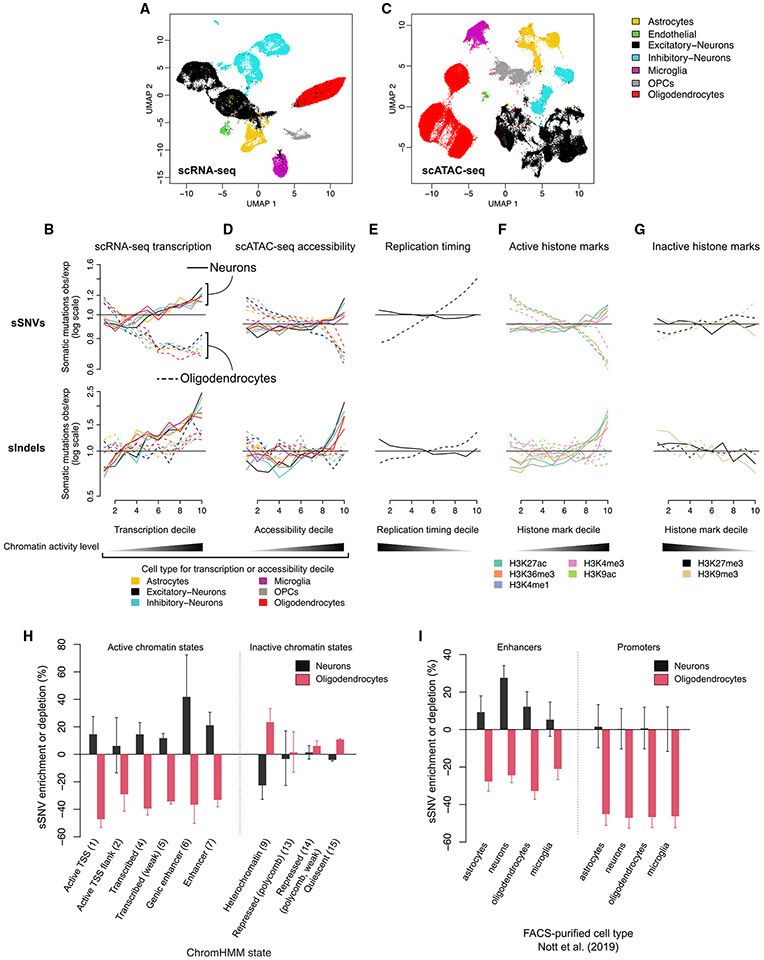
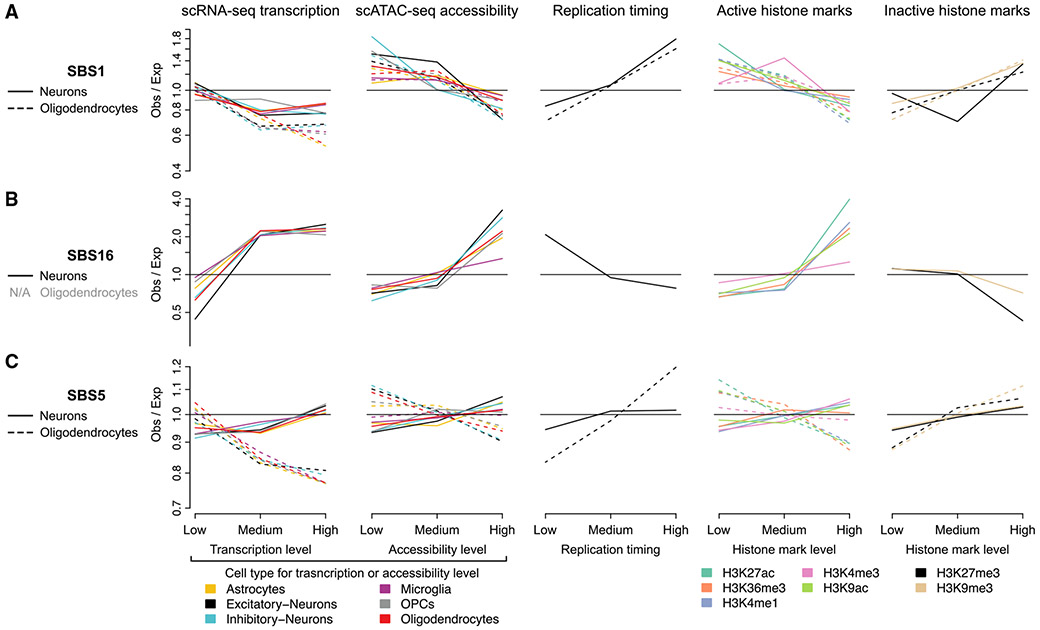
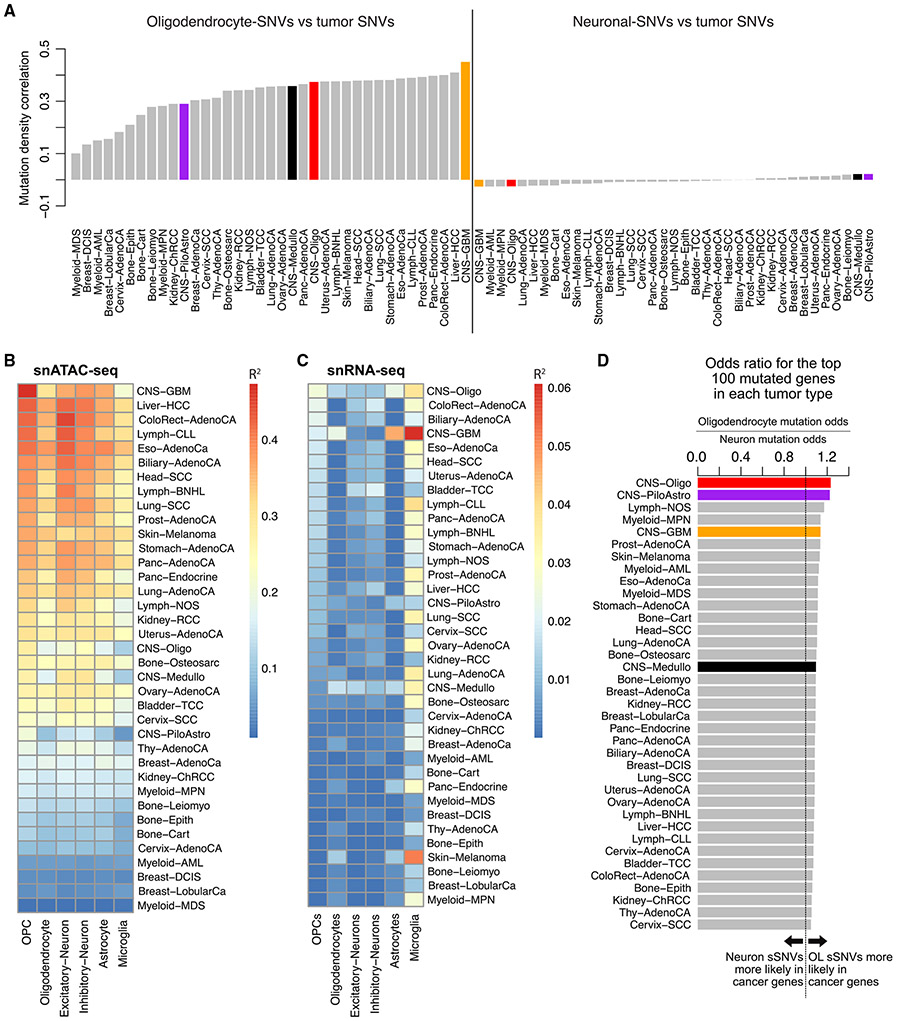
Characterizing somatic mutations in the brain is important for disentangling the complex mechanisms of aging, yet little is known about mutational patterns in different brain cell types. Here, we performed whole-genome sequencing (WGS) of 86 single oligodendrocytes, 20 mixed glia, and 56 single neurons from neurotypical individuals spanning 0.4-104 years of age and identified >92,000 somatic single-nucleotide variants (sSNVs) and small insertions/deletions (indels). Although both cell types accumulate somatic mutations linearly with age, oligodendrocytes accumulated sSNVs 81% faster than neurons and indels 28% slower than neurons. Correlation of mutations with single-nucleus RNA profiles and chromatin accessibility from the same brains revealed that oligodendrocyte mutations are enriched in inactive genomic regions and are distributed across the genome similarly to mutations in brain cancers. In contrast, neuronal mutations are enriched in open, transcriptionally active chromatin. These stark differences suggest an assortment of active mutagenic processes in oligodendrocytes and neurons.
Keywords: aging; brain cancer; brain disorders; glial cells; glioma; gliomagenesis; oligodendrocyte precursor cells; oligodendrocytes; somatic mutations.
Copyright © 2024 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests J.G. is now a Merck Research Laboratories (MRL) employee, and no work related to this manuscript was performed at MRL. P.J.P. is a member of the scientific advisory board for Bioskryb Genomics, Inc.
Figures
References
-
- Lee-Six H, Olafsson S, Ellis P, Osborne RJ, Sanders MA, Moore L, Georgakopoulos N, Torrente F, Noorani A, Goddard M, et al. (2019). The landscape of somatic mutation in normal colorectal epithelial cells. Nature 574, 532–537. - PubMed
MeSH terms
Substances
Grants and funding
- R01 HG012573/HG/NHGRI NIH HHS/United States
- P50 HD105351/HD/NICHD NIH HHS/United States
- R01 CA269805/CA/NCI NIH HHS/United States
- P30 AG072978/AG/NIA NIH HHS/United States
- U54 NS115266/NS/NINDS NIH HHS/United States
- U54 HD090255/HD/NICHD NIH HHS/United States
- R01 NS032457/NS/NINDS NIH HHS/United States
- DP2 AG086138/AG/NIA NIH HHS/United States
- UM1 DA058230/DA/NIDA NIH HHS/United States
- T32 HG002295/HG/NHGRI NIH HHS/United States
- R01 AG078929/AG/NIA NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- R01 AG082346/AG/NIA NIH HHS/United States
- K08 AG065502/AG/NIA NIH HHS/United States
- R01 AG070921/AG/NIA NIH HHS/United States
- P50 CA165962/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
